Двигательные единицы. Типы двигательных единиц. Понятие о двигательном аппарате. Виды и функции двигательных единиц (ДЕ). Композиция мышц Двигательные единицы тем меньше чем
Совокупность мотонейрона и иннервируемых им мышечных волокон называют двигательной (нейромоторной) единицей. Число мышечных волокон двигательной единицы варьирует в широких пределах в разных мышцах. Двигательные единицы невелики в мышцах, приспособленных для быстрых движений, от нескольких мышечных волокон до нескольких десятков их (мышцы пальцев, глаза, языка). Наоборот, в мышцах, осуществляющих медленные движения (поддержаниеттозы мышцами туловища), двигательные единицы велики и включают сотни и тысячи мышечных волокон.
При сокращении мышцы в натуральных (естественных) условиях можно зарегистрировать ее электрическую активность (элек-тромиограмму - ЭМГ) с помощью игольчатых или накожных электродов. В абсолютно расслабленной мышце электрическая активность почти отсутствует. При небольшом напряжении, например при поддержании позы, двигательные единицы разряжаются с небольшой частотой (5-10 имп/с), при большом напряжении частота импульсации повышается в среднем до 20-30 имп/с. ЭМГ позволяет судить о функциональной способности нейромоторных единиц. С функциональной точки зрения двигательные единицы разделяют на медленные и быстрые.
Медленные двигательные единицы включают медленные мотонейроны и медленные мышечные волокна (красные). Медленные мотонейроны, как правило, низкопороговые, так как обычно это малые мотонейроны. Устойчивый уровень импульсации у медленных мотонейронов наблюдается уже при очень слабых статических сокращениях мышц, при поддержании позы. Медленные мотонейроны способны поддерживать длительный разряд без заметного снижения частоты импульсации на протяжении длительного времени. Поэтому их называют малоутомляемыми или не-утомляемыми мотонейронами. В окружении медленных мышечных волокон богатая капиллярная сеть, позволяющая получать большое количество кислорода из крови. Повышенное содержание миоглобина облегчает транспорт кислорода в мышечных клетках к митохондриям. Миоглобин обусловливает красный цвет этих волокон. Кроме того, волокна содержат большое количество митохондрий и субстратов окисления - жиров. Все это обусловливает использование медленными мышечными волокнами более эффективного аэробного окислительного пути энергопродукции и определяет их высокую выносливость.
Быстрые двигательные единицы состоят из быстрых мотонейронов и быстрых мышечных волокон. Быстрые высокопороговые мотонейроны включаются в активность только для обеспечения относительно больших по силе статических и динамических сокращений мышц, а также в начале любых сокращений, чтобы увеличить скорость нарастания напряжения мышцы или сообщить движущейся части тела необходимое ускорение. Чем больше скорость и сила движений, т. е. чем больше мощность сократительного акта, тем больше участие быстрых двигательных единиц. Быстрые мотонейроны относятся к утомляемым - они не способны к длительному поддержанию высокочастотного разряда.
Быстрые мышечные волокна (белые мышечные волокна) более толстые, содержат больше миофибрилл, обладают большей силой, чем медленные волокна. Эти волокна окружает меньше капилляров, в клетках меньше митохондрий, миоглобина и жиров. Активность окислительных ферментов в быстрых волокнах ниже, чем в медленных, однако активность гликолитических ферментов, запа сы гликогена выше. Эти волокна не обладают большой выносливостью и более приспособлены для мощных, но относительно кратковременных сокращений. Активность быстрых волокон имеет значение для выполнения кратковременной высокоинтенсивной работы, например бега на короткие дистанции.
Выделяют также тонические мышечные волокна, они имеют 7-10 синапсов, принадлежащих, как правило, нескольким мотонейронам. ПКП этих мышечных волокон не вызывает генерацию ПД в них, а непосредственно запускает мышечное сокращение.
Скорость сокращения мышечных волокон находится в прямой зависимости от активности миозин-АТФ-азы - фермента, расщепляющего АТФ и тем самым способствующего образованию поперечных мостиков и взаимодействию актиновых и миозиновых мио-филаментов. Более высокая активность этого фермента в быстрых мышечных волокнах обеспечивает и более высокую скорость их сокращения по сравнению с медленными волокнами.
Движение - необходимое условие развития и существования организма, его приспособления к окружающей среде. Именно движение является основой целенаправленного поведения, что раскрывается словами Н.А.Бернштейна: «Очевидная огромная биологическая значимость двигательной деятельности организмов - почти единственной формы осуществления не только взаимодействия с окружающей средой, но и активного воздействия на эту среду, изменяющего ее с небезразличными для особи результатами…». Еще одним проявлением значимости движений является то, что в основе любой профессиональной деятельности лежит работа мышц.
Все многообразие двигательной деятельности осуществляется с помощью опорно-двигательного аппарата . Его составляют специализированные анатомические образования: мышцы, скелет и центральная нервная система.
В опорно-двигательном аппарате с определенной степенью условности выделяют пассивную часть - скелет и активную часть - мышцы.
К скелету относятся кости и их соединения (например, суставы).
Скелет служит опорой внутренним органам, местом прикрепления мышц, защищает внутренние органы от внешних механических повреждений. В костях скелета расположен костный мозг - орган кроветворения. В состав костей входит большое количество минеральных веществ (в наибольшей степени представлены кальций, натрий, магний, фосфор, хлор). Кость представляет собой динамичную живую ткань с высокой чувствительностью к различным регуляторным механизмам, к эндо - и экзогенным влияниям. Кость - не только орган опоры, но и важнейший участник минерального обмена (подробнее - в разделе Обмен веществ). Интегральным показателем метаболической активности костной ткани служат продолжающиеся в течение всей жизни процессы активной перестройки и обновления костных структур. Эти процессы, с одной стороны, являются важным механизмом поддержания минерального гомеостаза, с другой - обеспечивают структурную адаптацию кости к меняющимся условиям функционирования, что особо значимо в связи с регулярными занятиями физической культурой и спортом. В основе постоянно протекающих процессов костной перестройки лежит активность костных клеток - остеобластов и остеокластов.
Мышцы за счет способности сокращаться приводят в движение отдельные части тела, а также обеспечивают поддержание заданной позы. Мышечное сокращение сопровождается выработкой большого количества тепла, а значит, работающие мышцы участвуют в теплообразовании. Хорошо развитые мышцы являются прекрасной защитой внутренних органов, сосудов и нервов.
Кости и мышцы, как по массе, так и по объему составляют значительную часть всего организма, в их соотношении имеются существенные половые различия. Мышечная масса взрослого мужчины - от 35 до 50 % (в зависимости от того, насколько развиты мышцы) от общей массы тела, женщины - примерно 32-36 %. У спортсменов, специализирующихся в силовых видах спорта, мышечная масса может достигать 50-55%, а у культуристов – 60-70% общей массы тела. На долю костей приходится 18 % от массы тела у мужчин и 16 % у женщин.
У человека различают три вида мышц:
поперечнополосатые скелетные мышцы;
поперечнополосатая сердечная мышца;
гладкие мышцы внутренних органов, кожи, сосудов.
Гладкие мышцы делятся на тонические (не способны развивать «быстрые» сокращения, в сфинктерах полых органов) и фазно-тонические (которые делятся на обладающие автоматией , т.е. способностью к спонтанной генерации фазных сокращений. Примером могут быть мышцы органов ЖКТ и мочеточников, и не обладающие этим свойством – мышечный слой артерий, семенных протоков, мышца радужки глаза, они сокращаются под влиянием импульсов вегетативной нервной системы. Двигательная иннервация гладких мышц осуществляется отростками клеток вегетативной нервной системы, чувствительная - отростками клеток спинальных ганглиев. Как правило, сокращение гладкой мускулатуры не может быть вызвано произвольно, в регуляции ее сокращений не участвует кора мозга. Функция гладких мышц заключается в том, чтобы поддерживать длительное напряжение, при этом они затрачивают в 5 - 10 раз меньше АТФ, чем понадобилось бы для выполнения такой же задачи скелетной мышце.
Гладкие мышцы обеспечивают функцию полых органов , стенки которых они образуют. Благодаря гладким мышцам осуществляется изгнание содержимого из мочевого пузыря, кишок, желудка, желчного пузыря, матки. Гладкие мышцы обеспечивают сфинктерную функцию – создают условия для хранения определенного содержимого в полом органе (мочи в мочевом пузыре, плода в матке). Изменяя просвет кровеносных сосудов, гладкие мышцы адаптируют регионарный кровоток к местным потребностям в кислороде и питательных веществ, участвуют в регуляции дыхания за счет изменения просвета бронхиального дерева.
Скелетные мышцы являются активной частью опорно-двигательного аппарата, обеспечивая целенаправленную деятельность, в первую очередь за счет произвольных движений (подробнее особенности их строения и принципов работы рассмотрены ниже).
Виды мышечных волокон
Мышцы состоят из мышечных волокон, обладающих разной силой, скоростью и длительностью сокращения, а также утомляемостью. Ферменты в них обладают разной активностью и представлены в различных изомерных формах. Заметно различие в них содержания дыхательных ферментов – гликолитических и окислительных. По соотношению миофибрилл, митохондрий и миоглобина различают так называемые белые, красные и промежуточные волокна . По функциональным особенностям мышечные волокна делят на быстрые, медленные и промежуточные . Если по активности АТФазы мышечные волокна различаются довольно резко, то степень активности дыхательных ферментов варьирует весьма значительно, поэтому наряду с белыми и красными существуют и промежуточные волокна.
Наиболее явно мышечные волокна различаются особенностями молекулярной организации миозина. Среди различных его изоформ существуют две основных – «быстрая» и «медленная». При постановке гистохимических реакций их различают по АТФазной активности. С этими свойствами коррелирует и активность дыхательных ферментов. Обычно в быстрых волокнах (FF-волокна - быстро сокращающиеся, fast twitch fibres), преобладают гликолитические процессы, они более богаты гликогеном, в них меньше миоглобина, поэтому их называют также белыми. В медленных волокнах , обозначаемых как S (ST) волокна (slow twitch fibres), напротив, выше активность окислительных ферментов, они богаче миоглобином, выглядят более красными. Они включаются при нагрузках в пределах 20-25% от максимальной силы и отличаются хорошей выносливостью.
FT - волокна, обладающие по сравнению с красными волокнами небольшим содержанием миоглобина, характеризуются высокой сократительной скоростью и возможностью развивать большую силу. По сравнению с медленными волокнами они могут вдвое быстрее сокращаться и развить в 10 раз большую силу. FT-волокна, в свою очередь, подразделяются на FTO-и FTG-волокна. Существенные различия между перечисленными типами мышечных волокон определяется способом получения энергии (рис.2.1).
Рис. 2.1Различия энергообеспечения у мышечных волокон разных типов (по http://medi.ru/doc/g740203.htm).
Получение энергии в FTO-волокнах происходит так же, как и в ST-волокнах, преимущественно путем окислительного фосфорилирования. В связи с тем, что этот процесс разложения протекает относительно экономично (на каждую молекулу глюкозы при разложении мышечного гликогена для получения энергии накапливается 39 энергетических фосфатных соединений), FTO-волокна имеют также относительно высокую сопротивляемость утомляемости. Накопление энергии в FTG-волокнах происходит преимущественно путем гликолиза, т. е. глюкоза в отсутствии кислорода распадается до еще относительно богатого энергией лактата. В связи с тем, что этот процесс распада неэкономичен (на каждую молекулу глюкозы для получения энергии накапливается всего лишь 3 энергетических фосфатных соединения), FTG-волокна относительно быстро утомляются, но, тем не менее они способны развить большую силу и, как правило, включаются при субмаксимальных и максимальных мышечных сокращениях.
Двигательные единицы
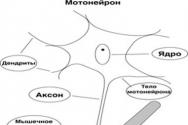
Основным морфофункциональным элементом нервно-мышечного аппарата скелетных мышц является двигательная единица – ДЕ (рис.2.2.).

Рис 2.2. Двигательная единица
ДЕ включает мотонейрон спинного мозга с иннервируемыми его аксоном мышечными волокнами. Внутри мышцы этот аксон образует несколько концевых веточек. Каждая такая веточка образует контакт – нервно-мышечный синапс на отдельном мышечном волокне. Нервные импульсы, идущие от мотонейрона, вызывают сокращения определенной группы мышечных волокон. ДЕ мелких мышц, осуществляющих тонкие движения (мышцы глаза, кисти), содержат небольшое количество мышечных волокон. В крупных мышцах их в сотни раз больше.
ДЕ активизируются по закону „все или ничего". Таким образом, если от тела мотонейрона переднего рога спинного мозга посылается по нервным путям импульс, то на него реагируют или все мышечные волокна ДЕ, или ни одного. Для бицепса это означает следующее: при нервном импульсе необходимой силы укорачиваются все сократительные элементы (миофибриллы) всех (примерно 1500) мышечных волокон соответствующей ДЕ.
Все ДЕ в зависимости от функциональных особенностей делятся на 3 группы:
I. Медленные неутомляемые . Они образованы "красными" мышечными волокнами, в которых меньше миофибрилл. Скорость сокращения и сила этих волокон относительно небольшие, но они мало утомляемы, поэтому эти волокна относят к тоническим. Регуляция сокращений таких, волокон осуществляется небольшим количеством мотонейронов, аксоны которых имеют мало концевых веточек. Пример – камбаловидная мышца.
II В. Быстрые, легко утомляемые . Мышечные волокна содержат много миофибрилл и называются "белыми". Быстро сокращаются и развивают большую силу, но быстро утомляются. Поэтому их называют фазными . Мотонейроны этих ДЕ самые крупные, имеют толстый аксон с многочисленными концевыми веточками. Они генерируют нервные импульсы большой частоты. Напр., мышцы глаза.
II А. Быстрые, устойчивые к утомлению (промежуточные).
Все мышечные волокна одной ДЕ относятся к одному и тому же типу волокон (FT- или ST-волокна).
Мышцы, задействованные в выполнении очень точных и дифференцированных движений (например, мышцы глаз или пальцев руки), состоят обычно из большого количества ДЕ (от 1500 до 3000). Такие ДЕ имеют небольшое количество мышечных волокон (от 8 до 50). Мышцы, выполняющие относительно менее точные движения (например, большие мышцы конечностей), обладают существенно меньшим количеством ДЕ, но в их состав включено большое число волокон (от 600 до 2000).
В среднем человек имеет примерно 40% медленных и 60 % быстрых волокон. Но это средняя величина (по всей скелетной мускулатуре), мышцы же выполняют различные функции. Количественный и качественный состав мышц неоднороден, в них входит различное число двигательных единиц, соотношение типов которых также различно (композиция мышц) . В связи с этим сократительные способности разных мышц неодинаковы. Наружные мышцы глаза, которые вращают глазное яблоко, развивают максимальное напряжение за одно сокращение длительностью всего 7.5 мс, камбалообразная – антигравитационная мышца нижней конечности, очень медленно развивает максимальное напряжение в течение 100 мс. Мышцы, выполняющие большую статическую работу (камбаловидная мышца), часто обладают большим количеством медленных ST-волокон, а мышцы, совершающие преимущественно динамические движения (бицепс), имеют большое количество FT-волокон.
Основные свойства мышечных волокон (следовательно, и двигательных единиц – ДЕ, в состав которых они входят), определяемые также свойствами мотонейронов, представлены в таблице 1.
text_fields
text_fields
arrow_upward
Мышечные волокна каждой Двигательной единицы (ДЕ) расположены на довольно значительном расстоянии друг от друга. Число мышечных волокон, входящих в одну ДЕ, отличается в разных мышцах. Оно меньше в мелких мышцах, осуществляющих тонкую и плавную регуляцию двигательной функции (например, мышцы кисти, глаза) и больше в крупных, не требующих столь точного контроля (икроножная мышца, мышцы спины). Так, в частности, в глазных мышцах одна ДЕ содержит 13-20 мышечных волокон, а ДЕ внутренней головки икроножной мышцы - 1500-2500.
Рис.4.8. Двигательные единицы (ДЕ) мышцы и их типы.
Мышечные волокна одной ДЕ имеют одинаковые морфофункциональные свойства.
По морфофункциональным свойствам ДЕ делятся на три основных типа (рис.4.8.):
I - медленные, неутомляемые;
II-А - быстрые, устойчивые к утомлению:
II-В - быстрые, легко утомляемые.
1 — медленные, слабые, неутомляемые мышечные волокна.
Низкий порог активации мотонейрона;
2 — промежуточный тип ДЕ;
3 — быстрые, сильные, быстроутомляемые мышечные
волокна. Высокий порог активаций мотонейрона.
Скелетные мышцы человека состоят из ДЕ всех трех типов. Одни из них включают преимущественно медленные ДЕ, другие - быстрые, третьи - и те, и другие.
Медленные, неутомляемые двигательные единицы (тип I)
text_fields
text_fields
arrow_upward
По сравнению с другими типами ДЕ у этих ДЕ наименьшие величины мотонейронов и, соответственно, наиболее низкие пороги их активации, меньшие толщина аксона и скорость проведения возбуждения по нему. Аксон разветвляется на небольшое число концевых веточек и иннервирует небольшую группу мышечных волокон. У мотонейронов медленных ДЕ сравнительно низкая частота разрядов (6-10 имп/с). Они начинают функционировать уже при малых мышечных усилиях. Так, мотонейроны ДЕ камбаловидной мышцы человека при удобном стоянии работают с частотой 4 имп/с. Устойчивая частота их импульсации составляет 6- 8 имп/с. С повышением силы сокращения мышцы частота разрядов мотонейронов медленных ДЕ повышается незначительно. Мотонейроны медленных ДЕ способны поддерживать постоянную частоту разрядов в течение десятков минут.
Мышечные волокна медленных ДЕ развивают небольшую силу при сокращении в связи с наличием в них меньшего, по сравнению с быстрыми волокнами, количества миофибрилл. Скорость сокращения этих волокон в 1,5-2 раза меньше, чем быстрых. Основными причинами этого являются низкая активность миозин АТФ-азы и меньшие скорость выхода ионов кальция из саркоплазматического ре-тикулума и его связывания с тропонином в процессе возбуждения волокна.
Мышечные волокна медленных ДЕ малоутомляемы. Они обладают хорошо развитой капиллярной сетью. На одно мышечное волокно, в среднем, приходится 4- 6 капилляров. Благодаря этому во время сокращения они обеспечиваются достаточным количеством кислорода. В их цитоплазме имеется большое количество митохондрий и высокая активность окислительных ферментов. Все это определяет существенную аэробную выносливость данных мышечных волокон и позволяет выполнять работу умеренной мощности длительное время без утомления.
Быстрые, легко утомляемые ДЕ (тип II-В)
text_fields
text_fields
arrow_upward
Из всех типов ДЕ мотонейроны этого типа наиболее крупные, имеют толстый аксон, разветвляющийся на большое число концевых веточек и иннервирующий соответственно большую группу мышечных волокон. По сравнению с другими эти мотонейроны обладают наиболее высоким порогом возбуждения, а их аксоны - большей скоростью проведения нервных импульсов.
Частота импульсации мотонейронов возрастает с ростом силы сокращения, достигая при максимальных напряжениях мышцы 25-50 имп/с. Эти мотонейроны не способны в течение длительного времени поддерживать устойчивую частоту разрядов, то есть быстро утомляются.
Мышечные волокна быстрых ДЕ, в отличие от медленных, содержат большее число сократительных элементов - миофибрилл, поэтому при сокращении развивают большую силу. Благодаря высокой активности миозиновой АТФ-азы у них выше скорость сокращения. Волокна этого типа содержат больше гликолитических ферментов, меньше митохондрий и миоглобина, окружены меньшим, по сравнению с медленными ДЕ, количеством капилляров. Эти волокна быстро утомляются. Более всего они приспособлены для выполнения кратковременной, но мощной работы (см. главу 27).
Быстрые, устойчивые к утомлению ДЕ (тип II-А)
text_fields
text_fields
arrow_upward
По своим морфофункциональным свойствам этот тип мышечных волокон занимает промежуточные положения между ДЕ I и II- В типов. Это сильные, быстро сокращающиеся волокна, обладающие большой аэробной выносливостью благодаря присущей им возможности использовать для получения энергии как аэробные, так и анаэробные процессы.
У разных людей соотношение числа медленных и быстрых ДЕ в одной и той же мышце определено генетически и может отличаться весьма значительно. Так, например, в четырехглавой мышце бедра человека процент медленных волокон может варьировать от 40 до 98%. Чем больше в мышце процент медленных волокон, тем более она приспособлена к работе на выносливость. И наоборот, лица с высоким процентом быстрых сильных волокон в большей мере способны к работе, требующей большой силы и скорости сокращения мышц.
Величина сокращения (сила мышцы) зависит от морфологических свойств и физиологического состояния мышцы:
1. Исходной длины мышцы (длинны покоя). Сила мышечного сокращения зависит от исходной длины мышцы или длины покоя. Чем сильнее мышца растянута в покое, тем сильнее сокращение (закон Франка-Старлинга).
2. Диаметра мышцы или поперечного сечения. Выделяют два диаметра:
а) анатомический диаметр - поперечное сечение мышц.
б) физиологический диаметр - перпендикулярное сечение каждого мышечного волокна. Чем больше физиологическое сечение, тем большей силой обладает мышца.
Сила мышцы измеряется весом максимального груза поднятого на высоту или максимальным напряжением, которое она способна развить в условиях изометрического сокращения. Измеряется в килограммах или ньютонах. Методика измерения силы мышцы называется динамометрия .
Выделяют два вида силы мышцы:
1. Абсолютная сила - отношение максимальной силы к физиологическому диаметру.
2. Относительная сила - отношение максимальной силы к анатомическому диаметру.
При сокращении мышца способна выполнять работу. Работа мышцы измеряется произведением поднятого груза на величину укорочения.
Работа мышцы характеризуется мощностью. Мощность мышцы определяется величиной работы в единицу времени и измеряется в ваттах.
Наибольшая работа и мощность достигается при средних нагрузках.
Мотонейрон с группой иннервируемых им мышечных волокон составляет двигательную единицу. Аксон мотонейронов может ветвиться и иннервировать группу мышечных волокон. Так, один аксон может иннервировать от 10 до 3000 мышечных волокон.
Различают двигательные единицы по строению и функциям.
По строению двигательные единицы делятся на:
1. Малые двигательные единицы, которые имеют малый мотонейрон и тонкий аксон, способный иннервировать 10-12 мышечных волокон. Например, мышцы лица, мышцы пальцев рук.
2. Большие двигательные единицы представлены крупным телом мотонейрона, толстым аксоном, который способен иннервировать более 1000 мышечных волокон. Например, четырехглавая мышца.
По функциональному значению двигательные единицы делятся на:
1. Медленные двигательные единицы. Они включают малые двигательные единицы, являются легко возбудимыми, характеризуются невысокой скоростью распространения возбуждения, в работу включаются первыми, но при этом они практически не утомляемы.
2. Быстрые двигательные единицы. Они состоят из больших двигательных единиц, плохо возбудимы, обладают большой скоростью проведения возбуждения. Обладают высокой силой и скоростью ответа. Например, мышцы боксера.
Эти особенности двигательных единиц обусловлены рядом свойств.
Мышечные волокна, которые входят в двигательные единицы, имеют сходные свойства и различия. Так, медленные мышечные волокна обладают:
1. Богатой капиллярной сетью.
4. В них содержится много жиров.
Благодаря этим особенностям эти мышечные волокна обладают высокой выносливостью, способны к небольшим по силе сокращениям, но длительным по времени.
Отличительные особенности быстрых мышечных волокон:
2. Обладают большей скоростью и силой сокращения.
В связи с этими особенностями быстрые мышечные волокна быстро утомляемы, но обладают большой силой и высокой скоростью ответа.
Двигательная единица включает двигательный нейрон вместе с иннервируемой им группой мышечных волокон. В разных мышцах двигательные единицы включают разное количество мышечных волокон. Так, в глазодвигательных мышцах на 1 нейрон приходится около 10 мышечных волокон, а в крупных мышцах туловища – больше 1000 волокон. Малые двигательные единицы обеспечивают быстрые и точные движения. Различают 3 типа двигательных единиц: быстрый, утомляемый; медленный, малоутомляемый; быстрый малоутомляемый. В любой мышце есть все типы волокон, но в разном соотношении. В мышцах спортсменов-спринтеров наблюдается больше быстрых мышечных волокон, а у стайеров – больше медленных мышечных волокон. Быстрые волокна хуже кровоснабжаются, поэтому способны к кратковременной работе. Медленные волокна обильно кровоснабжаются и могут длительно работать без утомления. Тела двигательных нейронов медленных двигательных единиц имеют небольшой размер и низкий порог возбудимости, т. е. могут быть активированы даже слабыми сигналами. Тела двигательных нейронов быстрых двигательных единиц более крупные, но менее возбудимые, они включаются, когда нужно развить большую силу.
Механизм передачи возбуждения в центральных синапсах, возбуждающие медиаторы, формирование возбуждающего постсинаптического потенциала (ВПСП). Значение хеморегулируемых и потенциалзависимых ионных каналов.
Механизм передачи возбуждения в синапсе . Медиаторы – это химические посредники передачи информации в синапсе с одного нейрона на другой. Выделение медиатора из пресинаптического окончания возможно только, если пресинаптическая мембрана будет деполяризована поступившими к нервному окончанию импульсами. В пресинаптической мембране есть каналы для ионов кальция, которые при отсутствии возбуждения закрыты. Ионы кальция играют определяющую роль в выделении медиатора. При деполяризации пресинаптической мембраны пришедшем сюда возбуждением кальциевые каналы открываются, кальций из синаптической щели поступает в пресинаптическое окончание, обеспечивает слияние медиаторных пузырьков с пресинаптической мембраной и выделение медиатора в синаптическую щель. Выделившийся в синаптическую щель медиатор перемещается к постсинаптической мембране, там он связывается со специфическими рецепторами, выполняющими одновременно роль ионных каналов. Образовавшийся комплекс «медиатор – рецептор» повышает проницаемость постсинаптической мембраны для определенных ионов, в результате меняется разность потенциалов на постсинаптической мембране и формируется постсинаптический потенциал. В зависимости от природы медиатора и характера связывающих его рецепторов постсинаптическая мембрана может быть деполяризована, что характерно для возбуждающих синапсов или гиперполяризована, что типично для тормозных синапсов. Возбуждающий постсинаптический потенциал (ВПСП) формируется на постсинаптической мембране в ответ на действие возбуждающих медиаторов. К таким медиаторам относят: ацетилхолин, норадреналин, дофамин, серотонин. Медиатор взаимодействует с рецепторами постсинаптической мембраны как ключ с замком, т. е для каждого медиатора существует определенный тип рецепторов. В результате взаимодействия медиатора с рецепторами постсинаптической мембраны открываются натриевые каналы (возможно участие и кальциевых каналов). Натрий поступает внутрь клетки через постсинаптическую мембрану и деполяризует ее. Возникшая разность потенциалов на постсинаптической мембране называется возбуждающим постсинаптическим потенциалом. Если его величина будет достаточной, то во внесинаптической части мембраны нейрона формируются потенциалы действия. Прекращение действия медиатора обусловлено удалением его из синаптической щели либо за счет обратного «захвата» структурами пресинаптического окончания, либо разрушения его специальными ферментами постсинаптической мембраны. В синапсах может развиваться процесс торможения, о чем будет сказано далее.
14. Торможение в ЦНС и его физиологическая роль. Учение И. М. Сеченова о центральном торможении. Тормозные медиаторы. Механизмы пре- и постсинаптического торможения .
Впервые о торможение как о процессе в центральной нервной системе высказался И. М. Сеченов (1863 год). Раздражая кристалликами поваренной соли область таламуса у лягушки, Сеченов отметил замедление двигательной реакции. Он пришел к выводу, что в центральной нервной системе развивается процесс торможения и соответственно есть тормозные центры. Этот вид торможения был назван Сеченовым центральным. Постсинаптическое торможение развивается, если тормозной нейрон формирует синапсы либо на дендритах, либо на теле возбуждающего нейрона. Синапсы имеют те же структурные элементы: пре-, постсинаптическую мембрану, синаптическую щель и медиаторы. Только в данном случае участвуют тормозные медиаторы: ГАМК, глицин, ацитилхолин и др. Медиаторы вызывают на постсинаптической мембране изменение проницаемости не для натрия, а либо для хлора, либо для калия через активацию соответствующих рецепторов и открытие хемозависимых ионных каналов. Если открываются каналы для ионов Cl - , он проходит через постсинаптическую мембрану внутрь и гиперполяризует её. В результате величина мембранного потенциала возрастает, а возбудимость снижается. Если в тормозном синапсе активируются каналы дляK + , то по градиенту он выходит на поверхность постсинаптической мембраны, которая также гиперполяризуется. Величина гиперполяризации называется тормозным постсинаптическим потенциалом (ТПСП), а вид торможения - постсинаптическим. Пресинаптическое торможение наблюдается в аксо-аксональных синапсах. Здесь аксон тормозного нейрона образует синапс на аксоне возбуждающего нейрона, еще до его синапса с другим нейроном. Поэтому торможение и называется пресинаптическим. Этот вид торможения блокирует прохождение возбуждение по аксону и имеет значение для фильтрации информации в сенсорных нейронах. Роль торможения в центральной нервной системе. Торможение обеспечивает: упорядоченность распространения возбуждения; согласованность во взаимодействии центров; защитную, охранительную роль от перевозбуждения. Важность торможения доказывают примеры: при столбняке или при отравлении стрихнином в нервной системе блокируются тормозные синапсы, поэтому возбуждение приобретает неупорядоченный характер, в результате развиваются мышечные судороги и наступает смерть. Торможение – это процесс возбуждения специализированных нейронов, приводящий к угнетению развития и распространения возбуждения. Важно помнить что торможение – это локальный, местный не распространяющийся процесс, в отличие от возбуждения.