Структурной единицей мышечной ткани является. Виды мышечных тканей и их особенности. Гладкая мышечная ткань эпидермального происхождения
Оно представляет собой вытянутое цилиндрическое образование с заостренными концами длиной от 1 до 40 мм (а по некоторым данным, до 120 мм), диаметром 0,1 мм.
Мышечное волокно окружено оболочкой – сарколеммой, в которой под электронным микроскопом отчетливо выделяются 2 листка: внутренний – типичная плазмолемма, а наружный представляет собой тонкую соединительнотканную пластинку – базальную пластинку.
В узкой щели между плазмолеммой и базальной пластинкой располагаются мелкие клетки – миосателлиты.
Таким образом, мышечное волокно является комплексным образованием и состоит из следующих основных структурных компонентов:
Миосимпласта;
Клеток-миосателлитов;
Базальной пластинки.
Базалъная пластинка образована тонкими коллагеновыми и ретикулярными волокнами, относится к опорному аппарату и выполняет вспомогательную функцию передачи сил сокращения на соединительнотканные элементы мышцы.
Клетки-миосателлиты являются камбиальными (ростковыми) элементами мышечных волокон и играют роль в процессах их физиологической и репаративной регенерации.
Миосимпласт является основным структурным компонентом мышечного волокна как по объему, так и по выполняемым функциям. Он образуется посредством слияния самостоятельных недифференцированных мышечных клеток – миобластов.
Миосимпласт можно рассматривать как вытянутую гигантскую многоядерную клетку, состоящую из большого числа ядер, цитоплазмы (саркоплазмы), плазмолеммы, включений, общих и специальных органелл. В миосимпласте содержится несколько тысяч (до 10 тыс.) продольно вытянутых светлых ядер, располагающихся на периферии под плазмолеммой. Вблизи ядер локализуются фрагменты слабовыраженной зернистой эндоплазматической сети, пластинчатого комплекса и небольшое число митохондрий. Центриоли в симпласте отсутствуют. В саркоплазме содержатся включения гликогена и миоглобина, аналога гемоглобина эритроцитов.
Отличительной особенностью миосимпласта является также наличие в нем специализированных органелл, к которым относятся:
Миофибриллы;
Саркоплазматическая сеть;
Канальцы Т-системы.
Миофибриллы – сократительные элементы миосимпласта – в большом количестве (до 1–2 тыс.) локализуются в центральной части саркоплазмы миосимпласта. Они объединяются в пучки, между которыми содержатся прослойки саркоплазмы. Между миофибриллами локализуется большое число митохондрий (саркосом). Каждая миофибрилла простирается продольно на протяжении всего миосимпласта и своими свободными концами прикрепляется к его плазмолемме у конических концов. Диаметр миофибриллы составляет 0,2–0,5 мкм.
Миофибриллы неоднородны по протяжению и подразделяются:
На темные (анизотропные), или А-диски, которые образованы более толстыми миофиламентами (10–12 нм), состоящими из белка миозина;
Светлые (изотропные), или I-диски, которые образованы тонкими миофиламентами (5–7 нм), состоящими из белка актина.
Темные и светлые диски всех миофибрилл располагаются на одном уровне и обусловливают поперечную исчерченность всего мышечного волокна.
Темные и светлые диски состоят из еще более тонких волоконец - протофибрилл, или миофиламентов.
Посередине I-диска поперечно актиновым миофиламентам проходит темная полоска – телофрагма, или Z-линия, посредине А-диска проходит менее выраженная М-линия, или мезофрагма.
Актиновые миофиламенты посередине I-диска скрепляются белками, составляющими Z-линию, свободными концами частично входят в А-диск между толстыми миофиламентами. При этом вокруг 1 миозинового филамента располагаются в актиновых.
При частичном сокращении миофибриллы актиновые миофиламенты как бы втягиваются в А-диск, и в нем образуется светлая зона, или Н-полоска, ограниченная свободными концами актиновых миофиламентов. Ширина Н-полоски зависит от степени сокращения миофибриллы.
Участок миофибриллы, расположенный между 2 Z-линиями, носит название саркомера и является структурно-функциональной единицей миофибриллы.
Саркомер включает в себя А-диск и расположенные по сторонам от него 2 половины 1-диска.
Следовательно, каждая миофибрилла представляет собой совокупность саркомеров.
Именно в саркомере осуществляется процесс сокращения.
Конечные саркомеры каждой миофибриллы прикрепляются к плазмолемме миосимпласта актиновыми миофиламентами.
Структурные элементы саркомера в расслабленном состоянии можно, выразить формулой
Z + 1/21 + 1/2А + М + 1/2А + 1/21 + Z.
Процесс сокращения осуществляется посредством взаимодействия актиновых и миозиновых филаментов и образования между ними актинмиозиновых мостиков, посредством которых происходит втягивание актиновых миофиламентов в А-диски – укорочение саркомера. Для развития этого процесса необходимы 3 условия.
Наличие энергии в виде АТФ;
Наличие ионов кальция; наличие биопотенциала.
АТФ образуется в саркосомах (митохондриях), в большом числе локализованных между миофибриллами.
Выполнение 2 последних условий осуществляется с помощью еще 2 специализированных органелл – саркоплазматической сети и Т-каналъцев.
Саркоплазматическая сеть представляет собой видоизмененную гладкую эндоплазматическую сеть и состоит из расширенных полостей и анастомозирующих канальцев, окружающих миофибриллы. Она подразделяется на фрагменты, окружающие отдельные саркомеры. Каждый фрагмент состоит из 2 терминальных цистерн, соединенных полыми анастомозируюшими канальцами – L-каналъцами. При этом терминальные цистерны охватывают саркомер в области I-дисков, а канальцы – в области А-дисков.
В терминальных цистернах и канальцах содержатся ионы кальция, которые при поступлении нервного импульса и достижении волны деполяризации мембран саркоплазматической сети выходят из цистерн и канальцев и распределяются между актиновыми и миозиновыми миофиламентами, инициируя их взаимодействие. После прекращения волны деполяризации ионы кальция устремляются обратно в терминальные цистерны и канальцы.
Таким образом, саркоплазматическая сеть не только является резервуаром для ионов кальция, но и играет роль кальциевого насоса.
Волна деполяризации передается на саркоплазматическую сеть от нервного окончания вначале по плазмолемме, а затем по Т-канальцам. Они не являются самостоятельными структурными элементами и представляют собой трубчатые выпячивания плазмолеммы в саркоплазму.
Проникая вглубь, Т-канальцы разветвляются и охватывают каждую миофибриллу в пределах 1 пучка строго на одном уровне, обычно на уровне Z-полоски или несколько медиальнее - в области соединения актиновых и миозиновых миофиламентов. Следовательно, к каждому саркомеру подходят и окружают его 2 Т-канальца.
По сторонам от каждого Т-канальца располагаются 2 терминальные цистерны саркоплазматической сети соседних саркомеров, которые вместе с Т-канальцами составляют триаду. Между стенкой Т-канальца и стенками терминальных цистерн имеются контакты, через которые волна деполяризации передается на мембраны цистерн и обусловливает выход из них ионов кальция и начало сокращения. Таким образом, функциональная роль Т-канальцев заключается в передаче биопотенциала с плазмолеммы на саркоплазматическую сеть.
Регенерация скелетной мышечной ткани, как и у других тканей, подразделяется на 2 типа – физиологическую и репаративную.
Физиологическая регенерация проявляется в форме гипертрофии мышечных волокон, что выражается в увеличении их толщины и даже длины, увеличении числа органелл, главным образом миофибрилл, а также нарастании числа ядер, что в конечном счете проявляется увеличением функциональной способности мышечного волокна. Радиоизотопным методом установлено, что увеличение числа ядер в мышечных волокнах в условиях гипертрофии достигается за счет деления клеток миосателлитов и последующего вхождения в миосимпласт дочерних клеток.
Увеличение числа миофибрилл осуществляется посредством синтеза актиновых и миозиновых белков свободными рибосомами и последующей сборки этих белков в актиновые и миозиновые миофиламенты параллельно с соответствующими филаментами саркомеров. В результате этого вначале происходит утолщение миофибрилл, а затем их расщепление и образование дочерних миофибрилл. Кроме того, возможно образование новых актиновых и миозиновых миофиламентов не параллельно, а встык предшествующим миофибриллам, чем достигается их удлинение.
Саркоплазматическая сеть и Т-канальцы в гипертрофирующемся волокне образуются за счет разрастания предшествующих элементов.
При определенных видах мышечной тренировки может формироваться Преимущественно красный тип мышечных волокон (у стайеров) или белый тип мышечных волокон (у спринтеров).
Возрастная гипертрофия мышечных волокон интенсивно проявляется с началом двигательной активности организма (1–2 года), что обусловлено прежде всего усилением нервной стимуляции.
В старческом возрасте, а также в условиях малой мышечной нагрузки
наступают атрофия специальных и общих органелл, истончение мышечных волокон и снижение их функциональной способности.
Репаративная регенерация развивается после повреждения мышечных волокон.
Способ регенерации зависит от величины дефекта:
При значительных повреждениях на протяжении мышечного волокна миосателлиты в области повреждения и в прилежащих участках растормаживаются, усиленно пролиферируют, а затем мигрируют в область дефекта мышечного волокна, где выстраиваются в цепочки, формируя миотрубку. Последующая дифференцировка миотрубки приводит к восполнению дефекта и восстановлению целостности мышечного волокна;
В условиях небольшого дефекта мышечного волокна на его концах за счет регенерации внутриклеточных органелл образуются мышечные
почки, которые растут навстречу друг другу, а затем сливаются, приводя к закрытию дефекта.
Репаративная регенераиия и восстановление целостности мышечных волокон могут осуществляться лишь в следующих случаях.
Во-первых, при сохраненной двигательной иннервации мышечныхволокон;
Во-вторых, если в область повреждения не попадают элементы соединительной ткани (фибробласты), – иначе на месте дефекта мышечного волокна развивается соединительнотканный рубец.
Советским ученым А.Н. Студитским доказана возможность амтотрансплантаиии скелетной мышечной ткани и даже целых мышц при соблюдении определенных условий:
· механическое измельчение мышечной ткани трансплантата с целью растормаживания клеток-сателлитов и последующей их пролиферации;
· помещение измельченной ткани в фасциальное ложе;
· подшивание двигательного нервного волокна к измельченному трансплантату;
· наличие сократительных движений мышц-антагонистов и синергистов.
2. Скелетные мышцы получают следующую иннервацию:
· двигательную (эфферентную);
· чувствительную (афферентную);
· трофическую (вегетативную).
Двигательную (эфферентную) иннервацию скелетные мышцы туловища и конечностей получают от мотонейронов передних рогов спинного мозга, а мышцы лица и головы – от двигательных нейронов определенных черепных нервов.
К каждому мышечному волокну подходит или ответвление от аксона мотонейрона, или же весь аксон. В мышцах, обеспечивающих тонкие координированные движения (мышцы кистей, предплечий, шеи), каждое мышечное волокно иннервируется 1 мотонейроном. В мышцах, обеспечивающих преимущественно поддержание позы, десятки и даже
сотни мышечных волокон получают двигательную иннервацию от 1 мотонейрона посредством разветвления его аксона.
Двигательное нервное волокно, подойдя к мышечному волокну, проникает под эндомизий и базальную пластинку и распадается на терминали, которые вместе с прилежащим специфическим участком миосимпласта образуют аксо-мышечный синапс или моторную бляшку. Под влиянием нервного импульса волна деполяризации с нервного окончания передается на плазмолемму миосимпласта, распространяется далее по Т-канальцам и в области триад передается на терминальные цистерны саркоплазматической сети, обусловливая выход ионов кальция и начало процесса сокращения мышечного волокна.
Чувствительная (афферентная) иннервация скелетных мышц осуществляется псевдоуниполярными нейронами спинальных ганглиев, посредством разнообразных рецепторных окончаний дендритов этих клеток.
Рецепторные окончания скелетных мыши можно разделить на 2 группы: специфические рецепторные приборы, характерные только для скелетных мышц:
Мышечное веретено;
Сухожильный орган Гольджи;
неспецифические рецепторные окончания кустиковидной или древовидной формы, распределяющиеся в рыхлой соединительной ткани:
Эндомизия;
Перимизия;
Эпимизия.
Мышечные веретена – довольно сложно устроенные инкапсулированные приборы. В каждой мышце содержится от нескольких единиц до нескольких десятков и даже сотен мышечных веретен. Каждое мышечное веретено содержит не только нервные элементы, но и 10–12 специфических мышечных волокон – интрафузальных, окруженных капсулой. Эти волокна располагаются параллельно сократительным мышечным волокнам (экстрафузальным) и получают не только чувствительную, но и специальную двигательную иннервацию. Мышечные веретена воспринимают раздражения как при растяжении данной мышцы, вызванном сокращением мышц-антагонистов, так и при ее сокращении.
Сухожильные органы представляют собой специализированные инкапсулированные рецепторы, включающие несколько сухожильных волокон, окруженных капсулой, среди которых распределяются терминальные ветвления дендрита псевдоуниполярного нейрона. При сокращении мышцы сухожильные волокна сближаются и сдавливают нервные окончания. Сухожильные органы воспринимают только степень сокращения данной мышцы. Посредством мышечных веретен и сухожильных органов при участии спинальных центров обеспечивается автоматизм движений (например, при ходьбе).
Трофическая (вегетативная) иннервация обеспечивается вегетативной нервной системой (ВНС) (ее симпатической частью) и осуществляется в основном опосредованно, посредством иннервации сосудов.
Скелетные мышцы богато снабжаются кровью. В рыхлой соединительной ткани перимизия в большом количестве содержатся артерии и вены, артериолы, венулы и артериоло-венулярные анастомозы. В эндомизии располагаются только капилляры, преимущественно узкие (4,5–7 мкм), которые и обеспечивают трофику мышечного волокна. Мышечное волокно вместе с окружающими его капиллярами и двигательным окончанием составляет мион.
В мышцах содержится большое количество артериоло-венулярных анастомозов, обеспечивающих адекватное кровоснабжение при различной мышечной активности.
Мышечные ткани (textus musculares) представляют группу разных по происхождению тканей животных и человека, обладающих общим свойством - сократимостью. Это свойство осуществляется этими тканями благодаря наличию в них специальных сократительных структур - миофиламентов Различают следующие основные виды мышечных тканей:
гладкую (неисчерченную) мышечную ткань и поперечнополосатые (исчерченные) мышечные ткани. Последние, в свою очередь, подразделяют на скелетную мышечную ткань и сердечную мышечную ткань. Свойством сократимости обладают также некоторые специализированные разновидности других тканей. К ним относят так называемую эпителиально-мышечную ткань (в потовых и слюнных железах) и нейроглиальную мышечную ткань (в радужной оболочке глаза) (таблица 9).
Гладкая (неисчерченная) мышечная ткань
Гладкая мышечная ткань (textus muscularis nonstriatus) развивается из мезенхимы. Она составляет двигательный аппарат внутренних органов, кровеносных и лимфатических сосудов. Ее сокращения имеют медленный, тонический характер. Структурной единицей гладкой мышечной ткани является клетка удлиненной веретенообразной формы - гладкий миоцит. Она покрыта плазмолеммой, к которой снаружи примыкает базальная мембрана и соединительнотканные волокна. Внутри клетки в ее центре, в миоплазме имеется вытянутой формы ядро, вокруг которого расположены митохондрии и другие органеллы.
В миоплазме миоцитов под электронным микроскопом обнаружены сократительные белковые нити - миофиламенты. Различают миофиламенты актиновые, миозиновые и промежуточные. Актиновые н миозиновые миофиламенты обеспечивают сам акт сокращения, а промежуточные предохраняют гладкие миоциты отих избыточного расширения при укорочении. Миофиламенты гладких миоцитов не образуют дисков, поэтому эти клетки не имеют поперечной исчерченности, и получили название гладких, неисчерченных. Гладкие миоциты хорошо регенерируют. Они делятся митозом, могут развиваться из малодифференцированных соединительнотканных клеток, способны к гипертрофии. Между клетками располагается опорная строма гладкой мышечной ткани - коллагеновые и эластические волокна, образующие плотные сети вокруг каждой клетки. Гладкие мышечные клетки синтезируют сами волокна этой стромы.
Поперечнополосатые (исчерченные) мышечные ткани
Как уже было сказано, в эту группу поперечнополосатых мышечных тканей включают скелетную и сердечную мышечные ткани. Эти ткани объединяют прежде всего по признаку поперечной исчерченности их специальных органелл - миофибрилл. Однако по своему происхождению, общему плану строения н функциональным особенностям, эти два вида поперечнополосатых мышечных тканей существенно отличаются.
Поперечнополосатая скелетная мышечная ткань
Скелетная мышечная ткань (textus muscularis striatus sceletalis) развивается из сегментированной мезодермы, точнее из ее центральных участков, получивших название миотомов. Структурно-функциональной единицей этой ткани являются многоядерные миосимпласты -поперечнополосатые мышечные волокна. С поверхности они покрыты сарколеммой - сложным образованием, состоящим из трехслойной плазмолеммы мышечного волокна, базальной мембраны и прилежащей к ней снаружи сети соединительнотканных волокон. Под базальной мембраной, прилегая к плазмолемме мышечного волокна, располагаются особые мышечные клетки - сателлиты. Внутри мышечного волокна, в его саркоплазме, по периферии, расположены многочисленные ядра, а в центре, вдоль волокна, находятся специальные органеллы - миофибриллы. Митохондрии н другие общие органеллы в мышечном волокне расположены вокруг ядер и вдоль миофибрилл. Под электронным микроскопом миофибриллы состоят из нитей - миофиламентов - актниовых, более тон ких (диаметром около 5-7 нм) и более толстых -миозиновых (диаметром около 10-20 нм).
Актиновые миофиламенты, содержащие белок актин, образуют изотропные диски (I). Это светлые, не обладающие двойным лучепреломлением диски. В центре дисковI проходитZ-линия - телофрагма. Эта линия делит дискI на два полудиска. В области Z-линий расположены так называемые триады. Триады состоят из трубчатых элементов - Т-трубочек, образованных вдавлением плазмолеммы внутрь мышечного волокна. По этим трубочкам нервный импульс поступает к миофибриллам. В каждой триаде одна Т-трубочка контактирует с двумя терминальными цистернами саркоплазматической сети, что обеспечивает выброс ионов кальция, необходимых для сократительного акта. В области Z-линий дискаI сходятся концы актиновых миофиламентов. Миозиновые миофиламенты, содержащие белок миозин, образуют анизотропные (А) темные диски, обладающие двойным лучепреломлением. В центре диска А проходит М-линия - мезофрагма. В М-линни сходятся концы миозиновых миофибрилл и обнаружена сеть канальцев саркоплазматической сети. Чередование в миофибриллах темных и светлых дисков придает мышечному волокну поперечную исчерченность. Структурной единицей миофибрилл является миомер (саркомер) - это участок миофибриллы между двумя Z-линиями. Его формула - А+2 1 / 2 I .
По современным представлениям в каждом мышечном волокне различают: сократительный аппарат, состоящий из мнофибрилл, включающих актиновые и миозиновые миофиламенты; трофический аппарат, в который входит саркоплазма с ядрами и органеллами; специальный мембранный аппарат триад; опорный аппарат, включающий сарколемму с эндомизием и мембранными структурами линий Z и М; и, наконец, нервный аппарат, представленный двигательными нервно-мышечными окончаниями - моторными бляшками и чувствительными нервными окончаниями - нервно-мышечными веретенами.
В скелетной мышечной ткани различают белые и красные мышечные волокна. Белые мышечные волокна содержат мало саркоплазмы и миоглобина и много мнофибрилл. На поперечном срезе в белых мышечных волокнах хорошо видны плотно расположенные миофибриллы. Они обеспечивают сильное, но непродолжительное сокращение. Красные мышечные волокна содержат много саркоплазмы и, следовательно, много миоглобина и мало миофибрилл. На поперечном срезе в таких мышечных волокнах миофибриллы расположены рыхло в виде групп, образуя многоугольники, получившие название полей Конгейма. Эти поля разделены друг от друг прослойками саркоплазмы. Красные мышечные волокна содержат много митохондрий, они способны к длительному сокращению. В каждой скелетной мышце, как органе, имеются и белые, и красные мышечные волокна. Однако их соотношение в разных мышечных группах неодинаково.
Каждое мышечное волокно окружено снаружи прослойкой рыхлой волокнистой соединительной ткани, получившей название эндомизия (endomysium). Группы мышечных волокон окружены перимизием (perimysium), а сама мышца - плотной соединительнотканной оболочкой -эпимизием (epimysium).
Поперечнополосатая скелетная мышечная ткань способна к регенерации. Сокращение мышечной ткани трактуется с позиции теории скольжения: актиновые миофиламенты вдвигаются, скользит между миозиновыми.
Сердечная мышечная ткань
Сердечная мышечная ткань (textus muscularis cardiacus) - это поперечнополосатая (исчерченная) мышечная ткань. Однако она имеет ряд существенных в своем строении отличий от скелетной мышечкой ткани. Развивается эта ткань из висцерального листка мезодермы, точнее, из так называемой миоэпикардиальной пластинки. Структурной единицей сердечной мышечной ткани являются поперечнополосатые клетки -сердечные миоциты или кардиомиоциты (miocyti cardiaci) с одним или двумя ядрами, расположенными в центре. По периферии цитоплазмы в кардиомиоцитах расположены миофибриллы, имеющие такое же строение, как и в скелетном мышечном волокне. Вокруг ядра и вдоль миофибрилл располагается большое количество митохоидрий (саркосом). Кардиомиоциты отделены друг от друга вставочными дисками (disci intercalati), образованными десмосомами и щелевыми контактами. Кардиомиоциты посредством этих дисков объединяются конец в конец в сердечные мышечные волокна, анастомозирующие между собой и сокращающиеся как единое целое. В сердечной мышечной ткани различают кардиомиоциты, - сократительные или типичные и проводящие или атипичные, составляющие проводящую систему сердца. Проводящие кардиомиоциты более крупные, содержат меньше миофибрилл и митохондрий. Их ядра часто расположены эксцентрично.1. Виды мышечной ткани
2. Поперечно-полосатая скелетная ткань
6. Гладкая мышечная ткань
1. Свойством сократимости обладают практически все виды клеток, благодаря наличию в их цитоплазме сократительного аппарата, представленного сетью тонких микрофиламентов (5-7 нм), состоящих из сократительных белков - актина, миозина, тропомиозина и других. За счет взаимодействия названных белков микрофиламентов осуществляются сократительные процессы и обеспечивается движение в цитоплазме гиалоплазмы, органелл, вакуолей, образование псевдоподий и инвагинаций плазмолеммы, а также процессы фаго- и пиноцитоза, экзоцитоза, деления и перемещения клеток. Содержание сократительных элементов, а, следовательно, и сократительные процессы неодинаково выражены в разных типах клеток. Наиболее выражены сократительные структуры в клетках, основной функцией которых является сокращение. Такие клетки или их производные образуют мышечные ткани, которые обеспечивают сократительные процессы в полых внутренних органах и сосудах, перемещение частей тела относительно друг друга, поддержание позы и перемещение организма в пространстве. Помимо движения при сокращении выделяется большое количество тепла, а, следовательно, мышечные ткани участвуют в терморегуляции организма. Мышечные ткани неодинаковы по строению, источникам происхождения и иннервации, по функциональным особенностям. Наконец, следует отметить, что любая разновидность мышечной ткани, помимо сократительных элементов (мышечных клеток и мышечных волокон) включает в себя клеточные элементы и волокна рыхлой волокнистой соединительной ткани и сосуды, которые обеспечивают трофику мышечных элементов, осуществляют передачу усилий сокращения мышечных элементов на скелет. Однако, функционально ведущими элементами мышечных тканей являются мышечные клетки или мышечные волокна.
Классификация мышечных тканей
· Гладкая (неисчерченная)- мезенхимная;
· специальная - нейрального происхождения и эпидермального происхождения;
· Поперечно-полосатая (исчерченная)- скелетная;
· сердечная.
Как видно из представленной классификации мышечная ткань подразделяется по строению на две основные группы - гладкую и поперечно-полосатую. Каждая из двух групп в свою очередь подразделяется на разновидности, как по источникам происхождения, так и по строению и функциональным особенностям. Гладкая мышечная ткань , входящая в состав внутренних органов и сосудов, развивается из мезенхимы. К специальным мышечным тканям нейрального происхождения относятся гладкомышечные клетки радужной оболочки, эпидермального происхождения - миоэпителиальные клетки слюнных, слезных, потовых и молочных желез.
Поперечно-полосатая мышечная ткань подразделяется на скелетную и сердечную. Обе эти разновидности развиваются из мезодермы, но из разных ее частей: скелетная - из миотомов сомитов, сердечная - из висцерального листка спланхнотома.
Каждая разновидность мышечной ткани имеет свою структурно-функциональную единицу. Структурно-функциональной единицей гладкой мышечной ткани внутренних органов и радужной оболочки является гладкомышечная клетка - миоцит; специальной мышечной ткани эпидермального происхождения - корзинчатый миоэпителиоцит ; сердечной мышечной ткани - кардиомиоцит ; скелетной мышечной ткани - мышечное волокно.
2. Поперечно-полосатая скелетная мышечная ткань
Структурно-функциональной единицей поперечно полосатой мышечной ткани является мышечное волокно . Оно представляет собой вытянутое цилиндрическое образование с заостренными концами длиной от 1 мм до 40 мм (а по некоторым данным до 120 мм), диаметром 0,1 мм. Мышечное волокно окружено оболочкой - сарколеммой, в которой под электронным микроскопом отчетливо выделяются два листка: внутренний - является типичной плазмолеммой, а наружный представляет собой тонкую соединительнотканную пластинку - базальную пластинку . В узкой щели между плазмолеммой и базальной пластинкой располагаются мелкие клетки - миосателлиты . Таким образом, мышечное волокно является комплексным образованием и состоит из следующих основных структурных компонентов :
· миосимпласта;
· клеток миосателиттов;
· базальной пластинки.
Базальная пластинка образована тонкими коллагеновыми и ретикулярными волокнами, относится к опорному аппарату и выполняет вспомогательную функцию передачи сил сокращения на соединительнотканные элементы мышцы.
Клетки миосателлиты являются камбиальными (ростковыми) элементами мышечных волокон и играют роль в процессах их физиологической и репаративной регенерации.
Миосимпласт является основным структурным компонентом мышечного волокна как по объему, так и по выполняемым функциям. Он образуется посредством слияния самостоятельных недифференцированных мышечных клеток - миобластов . Миосимпласт можно рассматривать как вытянутую гигантскую многоядерную клетку, состоящую из большого числа ядер, цитоплазмы (саркоплазмы), плазмолеммы, включений, общих и специальных органелл. В миосимпласте содержится несколько тысяч (до 10 000) продольно вытянутых светлых ядер, располагающихся на периферии под плазмолеммой. Вблизи ядер локализуются фрагменты слабовыраженной зернистой эндоплазматической сети, пластинчатого комплекса и небольшое число митохондрий. Центриоли в симпласте отсутствуют. В саркоплазме содержатся включения гликогена и миоглобина, аналога гемоглобина эритроцитов. Отличительной особенностью миосимпласта является также наличие в нем специализированных органелл , к которым относятся:
· миофибриллы;
· саркоплазматическая сеть;
· канальцы Т-системы.
Миофибриллы - сократительные элементы миосимпласта - в большом количестве (до 1000-2000) локализуются в центральной части саркоплазмы миосимпласта. Они объединяются в пучки, между которыми содержатся прослойки саркоплазмы. Между миофибриллами локализуется большое число митохондрий (саркосом). Каждая миофибрилла простирается продольно на протяжении всего миосимпласта и своими свободными концами прикрепляется к его плазмолемме у конических концов. Диаметр миофибриллы составляет 0,2-0,5 мкм. По своему строению миофибриллы неоднородны по протяжению и подразделяются на темные (анизотропные) или А-диски, и светлые (изотропные) или I-диски. Темные и светлые диски всех миофибрилл располагаются на одном уровне и обуславливают поперечную исчерченность всего мышечного волокна. Темные и светлые диски в свою очередь состоят из еще более тонких волоконец - протофибрилл или миофиламентов . Темные диски образованы более толстыми миофиламентами (10-12 нм), состоящими из белка миозина. Светлые диски образованы тонкими миофиламентами (5-7 нм), состоящими из белка актина. Посредине I-диска поперечно актиновым миофиламентам проходит темная полоска - телофрагма или Z-линия, посредине А-диска проходит менее выраженная М-линия или мезофрагма. Актиновые миофиламенты по средине I-диска скрепляются белками, составляющими Z-линию, свободными концами частично входит в А-диск между толстыми миофиламентами. При этом, вокруг одного миозинового филамента располагаются 6 актиновых. При частичном сокращении миофибриллы актиновые миофиламенты как бы втягиваются в А-диск и в нем образуется светлая зона или Н-полоска, ограниченная свободными концами актиновых миофиламентов. Ширина Н-полоски зависит от степени сокращения миофибриллы.
Участок миофибриллы, расположенный между двумя Z-линиями носит название саркомера и является структурно-функциональной единицей миофибриллы. Саркомер включает в себя А-диск и расположенные по сторонам от него две половины I-диска. Следовательно, каждая миофибрилла представляет собой совокупность саркомеров. Именно в саркомере осуществляется процесс сокращения. Следует отметить, что конечные саркомеры каждой миофибриллы прикрепляются к плазмолемме миосимпласта актиновыми миофиламентами. Структурные элементы саркомера в расслабленном состоянии можно выразить формулой :
Z+1/2I+1/2A+M+1/2A+1/2I+Z
Процесс сокращения осуществляется посредством взаимодействия актиновых и миозиновых филаментов и образования между ними актин-миозиновых мостиков , посредством которых происходит втягивание актиновых миофиламентов в А-диски укорочение саркомера. Для развития этого процесса необходимы три условия :
· наличие энергии в виде АТФ;
· наличие ионов кальция;
· наличие биопотенциала.
АТФ образуется в саркосомах (митохондриях) в большом числе локализованных между миофибриллами. Выполнение двух последних условий осуществляется с помощью еще двух специализированных органелл -саркоплазматической сети и Т-канальцев.
Саркоплазматическая сеть представляет собой видоизмененную гладкую эндоплазматическую сеть и состоит из расширенных полостей и анастомозирующих канальцев, окружающих миофибриллы. При этом саркоплазматическая сеть подразделяется на фрагменты, окружающие отдельные саркомеры. Каждый фрагмент состоит из двух терминальных цистерн, соединенных полыми анастомозирующими канальцами - L-канальцами. При этом терминальные цистерны охватывают саркомер в области I-дисков, а канальцы - в области А-диска. В терминальных цистернах и канальцах содержатся ионы кальция, которые при поступлении нервного импульса и достижении волны деполяризации мембран саркоплазматической сети, выходят из цистерн и канальцев и распределяются между актиновыми и миозиновыми миофиламентами, инициируя их взаимодействие. После прекращения волны деполяризации ионы кальция устремляются обратно в терминальные цистерны и канальцы. Таким образом, саркоплазматическая сеть является не только резервуаром для ионов кальция, но и играет роль кальциевого насоса.
Волна деполяризации передается на саркоплазматическую сеть от нервного окончания вначале по плазмолемме, а затем по Т-канальцам, которые не являются самостоятельными структурными элементами. Они представляют собой трубчатые выпячивания плазмолеммы в саркоплазму. Проникая вглубь, Т-канальцы разветвляются и охватывают каждую миофибриллу в пределах одного пучка строго на одном уровне, обычно на уровне Z-полоски или несколько медиальнее - в области соединения актиновых и миозиновых миофиламентов . Следовательно, к каждому саркомеру подходят и окружают его два Т-канальца. По сторонам от каждого Т-канальца располагаются две терминальные цистерны саркоплазматической сети соседних саркомеров, которые вместе с Т-канальцами составляют триаду. Между стенкой Т-канальца и стенками терминальных цистерн имеются контакты, через которые волна деполяризации передается на мембраны цистерн и обуславливает выход из них ионов кальция и начало сокращения. Таким образом, функциональная роль Т-канальцев заключается в передаче биопотенциала с плазмолеммы на саркоплазматическую сеть.
Для взаимодействия актиновых и миозиновых миофиламентов и последующего сокращения кроме ионов кальция необходима также энергия в виде АТФ, которая вырабатывается в саркосомах, в большом количестве располагающихся между миофибриллами.
Процесс взаимодействия актиновых и миозиновых филаментов упрощенно можно представить в следующем виде. Под влиянием ионов кальция стимулируется АТФ-азная активность миозина, что приводит к расщеплению АТФ, с образованием АДФ и энергии. Благодаря выделившейся энергии устанавливаются мостики между актином и миозином (а конкретнее, образуются мостики между головками белка миозина и определенными точками на актиновом филаменте)и за счет укорочения этих мостиков происходит подтягивание актиновых филаментов между миозиновыми. Затем эти связи распадаются (опять же с использованием энергии) и головки миозина образуют новые контакты с другими точками на актиновом филаменте, но расположенными дистальнее предыдущих. Так происходит постепенное втягивание актиновых филаментов между миозиновыми и укорочение саркомера . Степень этого сокращения зависит от концентрации ионов кальция вблизи миофиламентов и от содержания АТФ. После смерти организма АТФ в саркосомах не образуется, ее остатки расходуются на образование актин-миозиновых мостиков, а на распад уже не хватает, следствием чего наступает посмертное окоченение мышц, которое прекращается после аутолиза (распада) тканевых элементов.
При полном сокращении саркомера актиновые филаменты достигают М-полоски саркомера. При этом исчезают Н-полоски и I-диски, а формула саркомера может быть выражена в следующем виде:
Z+1/2IA+M+1/2AI+Z
При частичном сокращении формулу саркомера можно представить в следующем виде:
Z+1/nI+1/nIA+1/2H+M+1/2H+1/nAJ+1/nI+Z
Одновременное содружественное сокращение всех саркомеров каждой миофибриллы приводит к сокращению всего мышечного волокна. Крайние саркомеры каждой миофибриллы прикрепляются актиновыми миофиламентами к плазмолемме миосимпласта, которая на концах мышечного волокна имеет складчатый характер. При этом, на концах мышечного волокна базальная пластинка не заходит в складки плазмолеммы. Ее прободают тонкие коллагеновые и ретикулярные волокна, проникают в углубления складок плазмолеммы и прикрепляются в тех ее местах, к которым с внутренней стороны прикрепляются актиновые филаменты дистальных саркомеров. Благодаря этому создается прочная связь миосимпласта с волокнистыми структурами эндомизия. Коллагеновые и ретикулярные волокна концевых мышечных волокон, вместе с волокнистыми структурами эндомизия и перимизия в совокупности образуют сухожилия мышц, которые прикрепляются к определенным точкам скелета или вплетаются в сетчатый слой дермы в области лица. Благодаря сокращению мышц происходит перемещение частей или всего организма, а также изменение рельефа лица.
В мышечной ткани различают два основных типа мышечных волокон, между которыми имеются промежуточные, отличающиеся между собой прежде всего особенностями обменных процессов и функциональными свойствами и в меньшей степени - структурными особенностями.
Волокна I типа - красные мышечные волокна - характеризуются прежде всего высоким содержанием в саркоплазме миоглобина (что и придает им красный цвет), большим числом саркосом, высокой активностью в них сукцинатдегидрогеназы (СДГ), высокой активностью АТФ-азы медленного типа. Эти волокна обладают способностью медленного, но длительного тонического сокращения и малой утомляемостью.
Волокна II типа - белые мышечные волокна - характеризуются незначительным содержанием миоглобина, но высоким содержанием гликогена, высокой активностью фосфорилазы и АТФ-базы быстрого типа. Функционально характеризуются способностью быстрого, сильного, но непродолжительного сокращения. Между двумя крайними типами мышечных волокон находятся промежуточные, характеризующиеся различными сочетаниями названных включений и разной активностью перечисленных ферментов.
Мышца как орган состоит из мышечных волокон, волокнистой соединительной ткани, сосудов и нервов. Мышца - это анатомическое образование, основным и функционально ведущим структурным компонентом которого является мышечная ткань. Поэтому не следует рассматривать как синонимы понятия мышечная ткань и мышца.
Волокнистая соединительная ткань образует прослойки в мышце: эндомизий, перимизий и эпимизий, а также сухожилия. Эндомизий окружает каждое мышечное волокно, состоит из рыхлой волокнистой соединительной ткани и содержит кровеносные и лимфатические сосуды, в основном капилляры, посредством которых обеспечивается трофика волокна. Коллагеновые и ретикулярные волокна эндомизия проникают в базальную пластинку мышечного волокна, тесно с ним связаны и передают силы сокращения волокна на точки скелета. Перимизий окружает несколько мышечных волокон, собранных в пучки. В нем содержатся более крупные сосуды (артерии и вены, а также артериоло-венулярные анастомозы).
Эпимизий или фасция окружает всю мышцу, способствует функционированию мышцы, как органа. Любая мышца содержит все типы мышечных волокон в различном количественном соотношении. В мышцах, обеспечивающих поддержание позы, преобладают красные волокна, в мышцах, обеспечивающих движение пальцев и кистей, преобладают белые или переходные волокна. Характер мышечного волокна может меняться в зависимости от функциональной нагрузки и тренировки. Установлено, что биохимические, структурные и функциональные особенности мышечного волокна зависят от иннервации. Перекрестная пересадка эфферентных нервных волокон и их окончаний с красного волокна на белое и наоборот приводит к изменению обмена, а также структурных и функциональных особенностей в этих волокнах на противоположный тип.
3. Гистогенез и регенерация мышечной ткани
Из миотомов мезодермы в определенные участки мезенхимы выселяются малодифференцированные клетки - миобласты , часть из которых выстраивается в виде цепочки в стык друг к другу. В области контактов миобластов цитолеммы исчезает и образуется симпластическое образование - миотрубка, в которой ядра в виде цепочки располагаются в середине, а по периферии начинают дифференцироваться из миофиламентов миофибриллы. К миотрубке подрастают нервные волокна, образуя двигательные нервные окончания. Под влиянием эфферентной нервной импульсации начинается перестройка мышечной трубки в мышечное волокно: ядра перемещаются на периферию симпласта к плазмолемме, а миофибриллы занимают его центральную часть, из гладкой эндоплазматической сети развивается саркоплазматическая сеть, окружающая каждую миофибриллу на всем ее протяжении. Плазмолемма миосимпласта образует глубокие трубчатые впячивания - Т-канальцы. За счет деятельности зернистой эндоплазматической сети вначале миобластов, а затем и мышечных труб синтезируются и выделяются с помощью пластинчатого комплекса белки и полисахариды, из которых формируется базальная пластинка мышечного волокна.
Следует отметить, что при формировании миотрубки, а затем и дифференцировки мышечного волокна часть миобластов не входит в состав симпласта, а прилежит к нему, располагаясь под базальной пластинкой. Эти клетки носят название миосателлитов и играют важную роль в процессах физиологической и репаративной регенерации. Установлено, что закладка поперечно-полосатых скелетных мышечных волокон (миогенез) происходит только в эмбриональный период. В постнатальном периоде осуществляется их дальнейшая дифференцировка и гипертрофия, но количество мышечных волокон даже в условиях интенсивной тренировки не увеличивается.
Регенерация скелетной мышечной ткани
В мышечной, как в других тканях, различают два вида регенерации - физиологическую и репаративную. Физиологическая регенерация проявляется в форме гипертрофии мышечных волокон, что выражается в увеличении их толщины и даже длины, увеличение числа органелл, главным образом миофибрилл, а также нарастании числа ядер, что в конечном счете проявляется увеличением функциональной способности мышечного волокна. Радиоизотопным методом установлено, что увеличение числа ядер в мышечных волокнах в условиях гипертрофии достигается за счет деления клеток миосателлитов и последующего вхождения в миосимпласт дочерних клеток.
Увеличение числа миофибрилл осуществляется посредством синтеза актиновых и миозиновых белков свободными рибосомами и последующей сборки этих белков в актиновые и миозиновые миофиламенты параллельно с соответствующими филаментами саркомеров. В результате этого вначале происходит утолщение миофибрилл, а затем их расщепление и образование дочерних миофибрилл. Кроме того возможно образование новых актиновых и миозиновых миофиламентов не параллельно, а встык предшествующим миофибриллам, чем достигается их удлинение. Саркоплазматическая сеть и Т-канальцы в гипертрофирующемся волокне образуются за счет разрастания предшествующих элементов. При определенных видах мышечной тренировки может формироваться преимущественно красный тип мышечных волокон (у стайеров) или белый тип мышечных волокон (у спринтеров). Возрастная гипертрофия мышечных волокон интенсивно проявляется с началом двигательной активности организма (1-2 года), что обусловлено прежде всего усилением нервной стимуляции. В старческом возрасте, а также в условиях малой мышечной нагрузки наступает атрофия специальных и общих органелл, истончение мышечных волокон и снижение их функциональной способности.
Репаративная регенерация развивается после повреждения мышечных волокон. При этом способ регенерации зависит от величины дефекта. При значительных повреждениях на протяжении мышечного волокна миосателлиты в области повреждения и в прилежащих участках растормаживаются, усиленно пролиферируют, а затем мигрируют в область дефекта мышечного волокна, где выстраиваются в цепочки, формируя миотрубку. Последующая дифференцировка миотрубки приводит к восполнению дефекта и восстановлению целостности мышечного волокна. В условиях небольшого дефекта мышечного волокна на его концах, за счет регенерации внутриклеточных органелл, образуются мышечные почки , которые растут навстречу друг другу, а затем сливаются, приводя к закрытию дефекта. Однако, репаративная регенерация и восстановление целостности мышечных волокон могут осуществляться при определенных условиях: во-первых, при сохраненной двигательной иннервации мышечных волокон, во-вторых, если в область повреждения не попадают элементы соединительной ткани (фибробласты). Иначе на месте дефекта мышечного волокна развивается соединительно-тканный рубец.
Советским ученым А. Н. Студитским доказана возможность аутотрансплантации скелетной мышечной ткани и даже целых мышц при соблюдении определенных условий :
· механическое измельчение мышечной ткани трансплантата, с целью растормаживания клеток-сателлитов и последующей их пролиферации;
· помещение измельченной ткани в фасциальное ложе;
· подшивание двигательного нервного волокна к измельченному трансплантату;
· наличие сократительных движений мышц-антагонистов и синергистов.
4. Иннервация и кровоснабжение скелетных мышц
Скелетные мышцы получают двигательную, чувствительную и трофическую (вегетативную) иннервацию. Двигательную (эфферентную) иннервацию скелетные мышцы туловища и конечностей получают от мотонейронов передних рогов спинного мозга, а мышцы лица и головы - от двигательных нейронов определенных черепных нервов. При этом к каждому мышечному волокну подходит или ответвление от аксона мотонейрона, или же весь аксон. В мышцах, обеспечивающих тонкие координированные движения (мышцы кистей, предплечий, шеи), каждое мышечное волокно иннервируется одним мотонейроном. В мышцах, обеспечивающих преимущественно поддержание позы, десятки и даже сотни мышечных волокон получают двигательную иннервацию от одного мотонейрона, посредством разветвления его аксона.
Двигательное нервное волокно , подойдя к мышечному волокну, проникает под эндомизий и базальную пластинку и распадается на терминали, которые вместе с прилежащим специфическим участком миосимпласта образуют аксо-мышечный синапс или моторную бляшку. Под влиянием нервного импульса волна деполяризации с нервного окончания передается на плазмолемму миосимпласта, распространяется далее по Т-канальцам и в области триад передается на терминальные цистерны саркоплазматической сети, обуславливая выход ионов кальция и начало процесса сокращения мышечного волокна.
Чувствительная (афферентная) иннервация скелетных мышц осуществляется псевдоуниполярными нейронами спинальных ганглиев, посредством разнообразных рецепторных окончаний дендритов этих клеток. Рецепторные окончания скелетных мышц можно разделить на две группы:
I. специфические рецепторные приборы, характерные только для скелетных мышц:
· мышечное веретено;
· сухожильный орган Гольджи;
II. неспецифические рецепторные окончания кустиковидной или древовидной формы, распределяющиеся в рыхлой соединительной ткани эндомизия, перимизия и эпимизия.
Мышечные веретена - довольно сложно устроенные инкапсулированные приборы. В каждой мышце содержится от нескольких единиц до нескольких десятков и даже сотен мышечных веретен. Каждое мышечное веретено содержит не только нервные элементы, но также 10-12 специфических мышечных волокон - интрафузальных , окруженных капсулой. Эти волокна располагаются параллельно сократительным мышечным волокнам (экстрафузальным) и получают не только чувствительную, но и специальную двигательную иннервацию. Мышечные веретена воспринимают раздражения как при растяжении данной мышцы, вызванном сокращением мышц-антагонистов, так и при ее сокращении.
Сухожильные органы представляют собой специализированные инкапсулированные рецепторы, включающие несколько сухожильных волокон, окруженных капсулой, среди которых распределяются терминальные ветвления дендрита псевдоуниполярного нейрона. При сокращении мышцы сухожильные волокна сближаются и сдавливают нервные окончания. Сухожильные органы воспринимают только степень сокращения данной мышцы. Посредством мышечных веретен и сухожильных органов при участии спинальных центров обеспечивается автоматизм движений (например, при ходьбе).
Трофическая иннервация обеспечивается вегетативной нервной системой (ее симпатической частью) и осуществляется в основном опосредованно, посредством иннервации сосудов.
Скелетные мышцы богато снабжаются кровью. В рыхлой соединительной ткани перимизия в большом количестве содержатся артерии и вены, артериолы, венулы и артериоло-венулярные анастомозы. В эндомизии располагаются только капилляры, преимущественно узкие (4,5-7 мкм), которые и обеспечивают трофику мышечного волокна. Мышечное волокно, вместе с окружающими его капиллярами и двигательным окончанием составляют мион. В мышцах содержится большое количество артериоло-венулярных анастомозов, обеспечивающих адекватное кровоснабжение при различной мышечной активности.
5. Сердечная поперечно-полосатая мышечная ткань
Структурно-функциональной единицей является клетка -кардиомиоцит . По строению и функциям кардиомиоциты подразделяются на две основные группы :
· типичные или сократительные кардиомиоциты, образующие своей совокупностью миокард;
· атипичные кардиомиоциты, составляющие проводящую систему сердца и подразделяющиеся в свою очередь на три разновидности.
Сократительный кардиомиоцит представляет собой почти прямоугольную клетку 50-120 мкм в длину, шириной 15-20 мкм, в центре которой локализуется обычно одно ядро. Покрыт снаружи базальной пластинкой. В саркоплазме кардиомиоцита по периферии от ядра располагаются миофибриллы, а между ними и около ядра локализуются в большом количестве митохондрии. В отличие от скелетной мышечной ткани, миофибриллы кардиомиоцитов представляют собой не отдельные цилиндрические образования, а по существу сеть, состоящую из анастомозирующих миофибрилл, так как некоторые миофиламенты как бы отщепляются от одной миофибриллы и наискось продолжаются в другую. Кроме того, темные и светлые диски соседних миофибрилл не всегда располагаются на одном уровне, и потому поперечная исчерченность в кардиомиоцитах выражена не столь отчетливо, как в скелетных мышечных волокнах. Саркоплазматическая сеть, охватывающая миофибриллы, представлена расширенными анастомозирующими канальцами. Терминальные цистерны и триады отсутствуют. Т-канальцы имеются, но они короткие, широкие и образованы не только углублением плазмолеммы, но и базальной пластинки. Механизм сокращения в кардиомиоцитах практически не отличается от такового в скелетных мышечных волокнах.
Сократительные кардиомиоциты , соединяясь встык друг с другом, образуют функциональные мышечные волокна, между которыми имеются многочисленные анастомозы. Благодаря этому из отдельных кардиомиоцитов формируется сеть - функциональный синтиций . Наличие щелевидных контактов между кардиомиоцитами обеспечивает одновременное и содружественное их сокращение вначале в предсердиях, а затем и в желудочках.
Области контактов соседних кардиомиоцитов носят название вставочных дисков. Фактически, никаких дополнительных структур (диском между кардиомиоцитами нет. Вставочные диски - это места контактов цитолеммы соседних кардиомиоцитов, включающие в себя простые, десмосомные и щелевидные контакты. Обычно во вставочных дисках различают поперечный и продольный фрагменты. В области поперечных фрагментов имеются расширенные десмосомные соединения. В этих же местах с внутренней стороны плазмолемм прикрепляются актиновые филаменты саркомеров. В области продольных фрагментов локализуются щелевидные контакты. Посредством вставочных дисков обеспечивается как механическая, так и метаболическая (прежде всего ионная) связь кардиомиоцитов.
Сократительные кардиомиоциты предсердий и желудочков несколько отличаются между собой по морфологии и функциям. Так, кардиомиоциты предсердий в саркоплазме содержат меньше миофибрилл и митохондрий, в них почти не выражены Т-канальцы, а вместо них под плазмолеммой выявляются в большом числе везикулы и кавеолы - аналоги Т-канальцев. Кроме того, в саркоплазме предсердных кардиомиоцитов у полюсов ядер локализуются специфические предсердные гранулы, состоящие из гликопротеиновых комплексов. Выделяясь из кардиомиоцитов в кровь предсердий, эти вещества влияют на уровень давления крови в сердце и сосудах, а также препятствуют образованию тромбов в предсердиях. Следовательно, предсердные кардиомиоциты, кроме сократительной, обладают и секреторной функцией. В желудочковых кардиомиоцитах более выражены сократительные элементы, а секреторные гранулы отсутствуют.
Вторая разновидность кардиомиоцитов - атипичные кардиомиоциты образуют проводящую систему сердца , состоящую из:
· синусо-предсердный узел;
· предсердно-желудочковый узел;
· предсердно-желудочковый пучок (пучок Гиса)ствол, правую и левую ножки;
· концевые разветвления ножек - волокна Пункинье.
Атипичные кардиомиоциты обеспечивают генерирование биопотенциалов, их проведение и передачу на сократительные кардиомиоциты.
По своей морфологии атипичные кардиомиоциты отличаются от типичным рядом особенностей :
· они крупнее (длина 100 мкм, толщина 50 мкм);
· в цитоплазме содержимся мало миофибрилл, которые расположены неупорядочено и потому атипичные кардиомиоциты не имеют поперечной исчерченности;
· плазмолемма не образует Т-канальцев;
· во вставочных дисках между этими клетками отсутствуют десмосомы и щелевидные контакты.
Атипичные кардиомиоциты различных отделов проводящей системы отличаются между собой по структуре и функциям и подразделяются на три основные разновидности :
· Р-клетки (пейсмекеры) водители ритма (I типа);
· переходные клетки (II типа);
· клетки пучка Гиса и волокон Пуркинье (III тип).
Клетки I типа (Р-клетки) составляют основу синусо-предсердного узла, а также в небольшом количестве содержатся в атриовентрикулярном узле. Эти клетки способны самостоятельно генерировать с определенной частотой биопотенциалы и передавать их на переходные клетки (II типа), а последние передают импульсы на клетки III типа, от которых биопотенциалы передаются на сократительные кардиомиоциты.
Источники развития кардиомиоцитов - миоэпителиальные пластинки, представляющие собой определенные участки висцеральных листков спланхнотома, а конкретнееиз целомического эпителия этих участков.
Иннервация сердечной мышечной ткани
Биопотенциалы сократительные кардиомиоциты получают из двух источников:
· из проводящей системы сердца (прежде всего из синусо-предсердного узла);
· из вегетативной нервной системы (из ее симпатической и парасимпатической части).
Регенерация сердечной мышечной ткани
Кардиомиоциты регенерируют только по внутриклеточному типу. Пролиферации кардиомиоцитов не наблюдается. Камбиальные элементы в сердечной мышечной ткани отсутствуют. При поражении значительных участков миокарда (в частности, при инфаркте миокарда) восстановление дефекта происходит за счет разрастания соединительной ткани и образования рубцов (пластическая регенерация). Естественно, что сократительная функция в этих участках отсутствует. Поражение проводящей системы сопровождается нарушением ритма сердечных сокращений.
6. Гладкие мышечные ткани
Подавляющая часть гладкой мышечной ткани организма (внутренних органов и сосудов) имеет мезенхимальное происхождение.
Структурно-функциональной единицей гладкой мышечной ткани внутренних органов и сосудов является миоцит . Представляет собой чаще всего веретенообразную клетку (длиной 20-500 мкм, диаметром 5-8 мкм), покрытую снаружи базальной пластинкой, но встречаются и отростчатые миоциты. В центре располагается вытянутое ядро, по полюсам которого локализуются общие органеллы: зернистая эндоплазматическая сеть, пластинчатый комплекс, митохондрии, цитоцентр. В цитоплазме содержатся толстые (17 нм) миозиновые и тонкие (7 нм) актиновые миофиламенты, которые располагаются в основном параллельно друг другу вдоль оси миоцита и не образуют А и I диски, чем и объясняется отсутствие поперечной исчерченности миоцитов. В цитоплазме миоцитов и на внутренней поверхности плазмолеммы встречаются многочисленные плотные тельца, к которым прикрепляются актиновые, миозиновые, а так же промежуточные филаменты. Плазмолемма образует небольшие углубления - кавеолы, которые рассматриваются как аналоги Т-канальцев. Под плазмолеммой локализуются многочисленные везикулы, которые вместе с тонкими канальцами цитоплазмы являются элементами саркоплазматической сети.
Механизм сокращения в миоцитах в принципе сходен с сокращением саркомеров в миофибриллах в скелетных мышечных волокнах. Он осуществляется за счет взаимодействия и скольжения актиновых миофиламентов вдоль миозиновых. Для такого взаимодействия также необходимы энергия в виде АТФ, ионы кальция и наличие биопотенциала. Биопотенциалы поступают от эфферентных окончаний вегетативных нервных волокон непосредственно на миоциты или опосредованно от соседних клеток через щелевидные контакты и передаются через кавеолы на элементы саркоплазматической сети, обуславливая выход из них ионов кальция в саркоплазму. Под влиянием ионов кальция развиваются механизмы взаимодействия между актиновыми и миозиновыми филаментами, аналогичные тем, которые происходят в саркомерах скелетных мышечных волокон, в результате чего происходит скольжение названных миофиламентов и перемещение плотных телец в цитоплазме. В миоцитах, кроме актиновых и миозиновых филаментов, имеются еще промежуточные, которые одним концом прикрепляются к цитоплазматическим плотным тельцам, а другим - прикрепительным тельцам на плазмолемме и таким образом передают усилия взаимодействия актиновых и миозиновых филаментов на сарколемму миоцита, чем и достигается его укорочение.
Миоциты окружены снаружи рыхлой волокнистой соединительной тканью - эндомизием и связаны друг с другом боковыми поверхностями. При этом, в области тесного контакта соседних миоцитов базальные пластинки прерываются. Миоциты соприкасаются непосредственно плазмолеммами и в этих местах имеются щелевидные контакты, через которые осуществляется ионная связь и передача биопотенциала с одного миоцита на другой, что приводит к одновременному и содружественному их сокращению. Цепь миоцитов, объединенных механической и метаболической связью, составляет функциональное мышечное волокно. В эндомизии проходят кровеносные капилляры, обеспечивающие трофику миоцитов, а в прослойках соединительной ткани между пучками и слоями миоцитов в перимизии проходят более крупные сосуды и нервы, а также сосудистые и нервные сплетения.
Эфферентная иннервация гладкой мышечной ткани осуществляется вегетативной нервной системой. При этом, терминальные веточки аксонов эфферентных вегетативных нейронов, проходя по поверхности нескольких миоцитов, образуют на них небольшие варикозные утолщения, которые несколько прогибают плазмолемму и образуют мионевральные синапсы . При поступлении нервных импульсов в синаптическую щель выделяются медиаторы (ацетилхолин или норадреналин), и обуславливают деполяризацию мембран миоцитов и последующее их сокращение. Через щелевидные контакты биопотенциалы переходят из одного миоцита на другой, что сопровождается возбуждением и сокращением и тех гладкомышечных клеток, которые не содержат нервных окончаний. Возбуждение и сокращение миоцитов обычно продолжительны и обеспечивают тоническое сокращение гладкой мышечной ткани сосудов и полых внутренних органов, в том числе гладкомышечных сфинктеров. В этих органах содержатся и многочисленные рецепторные окончания в виде кустиков, деревцев или диффузных полей.
Регенерация гладкой мышечной ткани осуществляется несколькими способами:
· посредством внутриклеточной регенерациигипертрофии при усилении функциональной нагрузки;
· посредством митотического деления миоцитов при их повреждении (репаративная регенерация);
· посредством дифференцировки из камбиальных элементов - из адвентициальных клеток и миофибробластов.
7. Специальные гладкомышечные ткани
Нейрального происхождения развиваются из нейроэктодермы, из краев стенки глазного бокала, являющегося выпячиванием промежуточного мозга. Из этого источника развиваются миоциты, которые образуют две мышцы радужной оболочки глаза: мышцу суживающую зрачок и мышцу расширяющую зрачок. По своей морфологии миоциты радужной оболочки не отличаются от мезенхимных миоцитов, однако, отличаются по иннервации. Каждый миоцит получает вегетативную эфферентную иннервацию (мышца расширяющая зрачок - симпатическую, мышца суживающая зрачок -парасимпатическую). Благодаря этому, названные мышцы сокращаются быстро и координировано, в зависимости от мощности светового пучка.
Эпидермального происхождения развиваются из кожной эктодермы и представляют собой не типичные веретеновидные миоциты, а клетки звездчатой формы - миоэпителиальные клетки, располагающиеся в концевых отделах слюнных, молочных, слезных и потовых желез, снаружи от секреторных клеток. В своих отростках миоэпителиальные клетки содержат актиновые и миозиновые филаменты, благодаря взаимодействию которых отростки клеток сокращаются и способствуют выделению секрета из концевых отделов и мелких протоков названных желез в более крупные протоки. Эфферентную иннервацию получают также из вегетативного отдела нервной системы.
Внутренних органов, кожи, сосудов.
Скелетные мышцы совместно со скелетом составляют опорно-двигательную систему организма, которая обеспечивает поддержание позы и перемещение тела в пространстве. Кроме того, они выполняют защитную функцию, предохраняя внутренние органы от повреждений.
Скелетные мышцы являются активной частью опорно-двигательного аппарата, включающего также кости и их сочленения, связки, сухожилия. Масса мышц может достигать 50% общей массы тела.
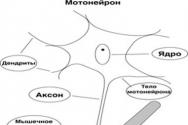
С функциональной точки зрения к двигательному аппарату можно отнести и моторные нейроны, посылающие нервные импульсы к мышечным волокнам. Тела моторных нейронов, иннервирующих аксонами скелетную мускулатуру, располагаются в передних рогах спинного мозга, а иннервирующих мышцы челюстно-лицевой области — в моторных ядрах ствола мозга. Аксон мотонейрона при входе в скелетную мышцу ветвится, и каждая веточка участвует в формировании нервно-мышечного синапса на отдельном мышечном волокне (рис. 1).
Рис. 1. Разветвления аксона моторного нейрона на аксонные терминалы. Электронограмма

Рис. Строение скелетной мышцы человека
Скелетные мышцы состоят из мышечных волокон, которые объединяются в мышечные пучки. Совокупность мышечных волокон, иннервируемых веточками аксона одного моторного нейрона, называют двигательной (или моторной) единицей. В глазных мышцах 1 двигательная единица может содержать 3-5 мышечных волокон, в мышцах туловища — сотни волокон, в камбаловидной мышце — 1500-2500 волокон. Мышечные волокна 1 двигательной единицы имеют одинаковые морфофункциональные свойства.
Функциями скелетных мышц являются:
- передвижение тела в пространстве;
- перемещение частей тела относительно друг друга, в том числе осуществление дыхательных движений, обеспечивающих вентиляцию легких;
- поддержание положения и позы тела.
Скелетные мышцы вместе со скелетом составляют опорно-двигательную систему организма, которая обеспечивает поддержание позы и перемещение тела в пространстве. Наряду с этим скелетные мышцы и скелет выполняют защитную функцию, предохраняя внутренние органы от повреждения.
Кроме того, поперечно-полосатые мышцы имеют значение в выработке тепла, поддерживающего температурный гомеостаз, и в депонировании некоторых питательных веществ.

Рис. 2. Функции скелетных мышц
Физиологические свойства скелетных мышц
Скелетные мышцы обладают следующими физиологическими свойствами.
Возбудимость. Обеспечивается свойством плазматической мембраны (сарколеммы) отвечать возбуждением на поступление нервного импульса. Из-за большей разности потенциала покоя мембраны поперечно-полосатых мышечных волокон (Е 0 около 90 мВ) возбудимость их ниже, чем нервных волокон (Е 0 около 70 мВ). Амплитуда потенциала действия у них больше (около 120 мВ), чем у других возбудимых клеток.
Это позволяет на практике достаточно легко регистрировать биоэлектрическую активность скелетных мыши. Длительность потенциала действия составляет 3-5 мс, что определяет короткую продолжительность фазы абсолютной рефрактерности возбужденной мембраны мышечных волокон.
Проводимость. Обеспечивается свойством плазматической мембраны формировать локальные круговые токи, генерировать и проводить потенциал действия. В результате потенциал действия распространяются по мембране вдоль мышечного волокна и вглубь по поперечным трубочкам, формируемым мембраной. Скорость проведения потенциала действия составляет 3-5 м/с.
Сократимость. Представляет собой специфическое свойство мышечных волокон изменять свою длину и напряжение вслед за возбуждением мембраны. Сократимость обеспечивается специализированными сократительными белками мышечного волокна.
Скелетные мышцы обладают также вязкоэластическими свойствами, имеющими важное значение для расслабления мышц.

Рис. Скелетные мышцы человека
Физические свойства скелетных мышц
Скелетные мышцы характеризуются растяжимостью, эластичностью, силой и способностью совершать работу.
Растяжимость - способность мышцы изменять длину под действием растягивающей силы.
Эластичность - способность мышцы восстанавливать первоначальную форму после прекращения действия растягивающей или деформирующей силы.
- способность мышцы поднимать груз. Для сравнения силы различных мышц определяют их удельную силу путем деления максимальной массы на число квадратных сантиметров ее физиологического сечения. Сила скелетной мышцы зависит от многих факторов. Например, от числа двигательных единиц, возбуждаемых в данный момент времени. Также она зависит от синхронности работы двигательных единиц. Сила мышцы зависит и от исходной длины. Существует определенная средняя длина, при которой мышца развивает максимальное сокращение.
Сила гладких мышц тоже зависит от исходной длины, синхронности возбуждения мышечного комплекса, а также от концентрации ионов кальция внутри клетки.
Способность мышцы совершать работу. Работа мышцы определяется произведением массы поднятого груза на высоту подъема.
Работа мышц возрастаете увеличением массы поднимаемого груза, но до определенного предела, после которого увеличение груза приводит к уменьшению работы, т.е. снижается высота подъема. Максимальная работа совершается мышцей при средних нагрузках. Это называется законом средних нагрузок. Величина мышечной работы зависит от числа мышечных волокон. Чем толще мышца, тем больший груз она может поднять. Длительное напряжение мышцы приводит к ее утомлению. Это обусловлено истощением энергетических запасов в мышце (АТФ, гликоген, глюкоза), накоплением молочной кислоты и других метаболитов.
Вспомогательные свойства скелетной мускулатуры
Растяжимость — это способность мышцы изменять свою длину под действием растягивающей ее силы. Эластичность — способность мышцы принимать свою первоначальную длину после прекращения действия растягивающей или деформирующей силы. Живая мышца обладает малой, но совершенной эластичностью: уже небольшая сила способна вызвать относительно большое удлинение мышцы, а возвращение ее к первоначальным размерам является полным. Это свойство очень важно для осуществления нормальных функций скелетных мышц.
Сила мышцы определяется максимальным грузом, который мышца в состоянии поднять. Для сравнения силы различных мышц определяют их удельную силу, т.е. максимальный груз, который мышца в состоянии поднять, делят на число квадратных сантиметров ее физиологического поперечного сечения.
Способность мышцы совершать работу. Работа мышцы определяется произведением величины поднятого груза на высоту подъема. Работа мышцы постепенно увеличивается с увеличением груза, но до определенного предела, после которого увеличение груза приводит к уменьшению работы, так как снижается высота подъема груза. Следовательно, максимальная работа мышцей производится при средних величинах нагрузок.
Утомление мышц. Мышцы не могут работать беспрерывно. Длительная работа приводит к снижению их работоспособности. Временное понижение работоспособности мышцы, наступающее при длительной работе и исчезающее после отдыха, называется утомлением мышцы. Принято различать два вида утомления мышц: ложное и истинное. При ложном утомлении утомляется не мышца, а особый механизм передачи импульсов с нерва на мышцу, называемый синапсом. В синапсе истощаются резервы медиаторов. При истинном утомлении в мышце происходят следующие процессы: накопление недоокисленных продуктов распада питательных веществ вследствие недостаточного поступления кислорода, истощение запасов источников энергии, необходимой для мышечного сокращения. Утомление проявляется уменьшением силы сокращения мышцы и степени расслабления мышцы. Если мышца на некоторое время прекращает работу и находится в состоянии покоя, то восстанавливается работа синапса, а с кровью удаляются продукты обмена и доставляются питательные вещества. Таким образом, мышца вновь приобретает способность сокращаться и производить работу.
Одиночное сокращение
Раздражение мышцы или иннервирующего ее двигательного нерва одиночным стимулом вызывает одиночное сокращение мышцы. Различают три основные фазы такого сокращения: латентная фаза, фаза укорочения и фаза расслабления.
Амплитуда одиночного сокращения изолированного мышечного волокна от силы раздражения не зависит, т.е. подчиняется закону «все или ничего». Однако сокращение целой мышцы, состоящей из множества волокон, при ее прямом раздражении зависит от силы раздражения. При пороговой силе тока в реакцию вовлекается лишь небольшое число волокон, поэтому сокращение мышцы едва заметно. С увеличением силы раздражения число волокон, охваченных возбуждением, возрастает; сокращение усиливается до тех пор, пока все волокна не оказываются сокращенными («максимальное сокращение») — этот эффект называется лестницей Боудича. Дальнейшее усиление раздражающего тока на сокращение мышцы не влияет.

Рис. 3. Одиночное сокращение мышцы: А — момент раздражения мышцы; а-6 — скрытый период; 6-в — сокращение (укорочение); в-г — расслабление; г-д — последовательные эластические колебания.
Тетанус мышцы
В естественных условиях к скелетной мышце из центральной нервной системы поступают не одиночные импульсы возбуждения, которые служат для нее адекватными раздражителями, а серии импульсов, на которые мышца отвечает длительным сокращением. Длительное сокращение мышцы, возникающее в ответ на ритмическое раздражение, получило название тетанического сокращения, или тетануса. Различают два вида тетануса: зубчатый и гладкий (рис. 4).
Гладкий тетанус возникает, когда каждый последующий импульс возбуждения поступает в фазу укорочения, а зубчатый - в фазу расслабления.
Амплитуда тетанического сокращения превышает амплитуду одиночного сокращения. Академик Н.Е. Введенский обосновал изменчивость амплитуды тетануса неодинаковой величиной возбудимости мышцы и ввел в физиологию понятия оптимума и пессимума частоты раздражения.
Оптимальной называется такая частота раздражения, при которой каждое последующее раздражение поступает в фазу повышенной возбудимости мышцы. При этом развивается тетанус максимальной величины (оптимальный).
Пессимальной называется такая частота раздражения, при которой каждое последующее раздражение осуществляется в фазу пониженной возбудимости мышцы. Величина тетануса при этом будет минимальной (пессимальной).

Рис. 4. Сокращение скелетной мышцы при разной частоте раздражения: I — сокращение мышцы; II — отметка частоты раздражения; а — одиночные сокращения; б- зубчатый тетанус; в — гладкий тетанус
Режимы мышечных сокращений
Для скелетных мышц характерны изотонический, изометрический и смешанный режимы сокращения.
При изотоническом сокращении мышцы изменяется ее длина, а напряжение остается постоянным. Такое сокращение происходит в том случае, когда мышца не преодолевает сопротивления (например, не перемещает груз). В естественных условиях близкими к изотоническому типу сокращениями являются сокращения мышц языка.
При изометрическом сокращении в мышце во время ее активности нарастает напряжение, но из-за того, что оба конца мышцы фиксированы (например, мышца пытается поднять большой груз), она не укорачивается. Длина мышечных волокон остается постоянной, меняется лишь степень их напряжения.
Сокращаются по аналогичным механизмам.
В организме сокращения мышц никогда не бывают чисто изотоническими или изометрическими. Они всегда имеют смешанный характер, т.е. происходит одновременное изменение и длины, и напряжения мышцы. Такой режим сокращения называется ауксотоническим, если преобладает напряжение мышцы, или ауксометрическим, если преобладает укорочение.
Существует три типа мышечной ткани и соответственно мышц, различающихся по структуре мышечных волокон и характеру иннервации:
1. Скелетная (поперечно-полосатая) мышечная ткань
2. Сердечная поперечно - полосатой мышечная ткань
3. Гладкая мышечная ткань
Скелетная (поперечно-полосатая) мышечная ткань
Упругая, эластичная ткань, способная сокращаться под влиянием нервных импульсов; один из типов мышечной ткани. Образует скелетную мускулатуру человека и животных, предназначенную для выполнения различных действий: движения тела, сокращения голосовых связок, дыхания.
Состоит из миоцитов, имеющих большую длину (до нескольких сантиметров) диаметром от 50 до 100 мкм. Клетки многоядерные, содержат до 100 и более ядер. Микроскопическое изучение показало, что волокно скелетной мышцы по всей длине имеет регулярную поперечную исчерченность в виде чередующихся светлых и темных участков, (поперечнополосатая мышечная ткань образована мышечными клетками, содержащими миофибриллы, которые состоят из миозиновых и актиновых протофибрилл, взаимное положение которых создает поперечную исчерченность),что послужило основанием для другого названия – поперечнополосатые мышцы.
Функции скелетных мышц находятся под контролем центральной нервной системы, т.е. контролируются нашей волей, поэтому их называют также произвольными мышцами. Однако они могут находиться в состоянии частичного сокращения и независимо от нашего сознания; такое состояние называют тонусом. мышечный ткань волокно
Сердечная поперечно - полосатой мышечная ткань
Структурно-функциональной единицей сердечной поперечно - полосатой мышечной ткани является клетка - кардиомиоцит. По строению и функциям кардиомиоциты подразделяются на две основные группы:
Типичные или сократительные кардиомиоциты, образующие своей совокупностью миокард;
Атипичные кардиомиоциты, составляющие проводящую систему сердца и подразделяющиеся в свою очередь на три разновидности.
Сократительный кардиомиоцит представляет собой почти прямоугольную клетку 50-120 мкм в длину, шириной 15-20 мкм, в центре которой локализуется обычно одно ядро. Покрыт снаружи базальной пластинкой. В саркоплазме кардиомиоцита по периферии от ядра располагаются миофибриллы, а между ними и около ядра локализуются в большом количестве митохондрии. В отличие от скелетной мышечной ткани, миофибриллы кардиомиоцитов представляют собой не отдельные цилиндрические образования, а по существу сеть, состоящую из анастомозирующих миофибрилл, так как некоторые миофиламенты как бы отщепляются от одной миофибриллы и наискось продолжаются в другую. Кроме того, темные и светлые диски соседних миофибрилл не всегда располагаются на одном уровне, и потому поперечная исчерченность в кардиомиоцитах выражена не столь отчетливо, как в скелетных мышечных волокнах. Саркоплазматическая сеть, охватывающая миофибриллы, представлена расширенными анастомозирующими канальцами. Терминальные цистерны и триады отсутствуют. Т-канальцы имеются, но они короткие, широкие и образованы не только углублением плазмолеммы, но и базальной пластинки. Механизм сокращения в кардиомиоцитах практически не отличается от такового в скелетных мышечных волокнах.
Сократительные кардиомиоциты, соединяясь встык друг с другом, образуют функциональные мышечные волокна, между которыми имеются многочисленные анастомозы. Благодаря этому из отдельных кардиомиоцитов формируется сеть - функциональный синтиций. Наличие щелевидных контактов между кардиомиоцитами обеспечивает одновременное и содружественное их сокращение вначале в предсердиях, а затем и в желудочках.
Области контактов соседних кардиомиоцитов носят название вставочных дисков. Фактически, никаких дополнительных структур (дисков) между кардиомиоцитами нет. Вставочные диски - это места контактов цитолеммы соседних кардиомиоцитов, включающие в себя простые, десмосомные и щелевидные контакты. Обычно во вставочных дисках различают поперечный и продольный фрагменты. В области поперечных фрагментов имеются расширенные десмосомные соединения. В этих же местах с внутренней стороны плазмолемм прикрепляются актиновые филаменты саркомеров. В области продольных фрагментов локализуются щелевидные контакты. Посредством вставочных дисков обеспечивается как механическая, так и метаболическая (прежде всего ионная) связь кардиомиоцитов.
Сократительные кардиомиоциты предсердий и желудочков несколько отличаются между собой по морфологии и функциям. Так, кардиомиоциты предсердий в саркоплазме содержат меньше миофибрилл и митохондрий, в них почти не выражены Т-канальцы, а вместо них под плазмолеммой выявляются в большом числе везикулы и кавеолы - аналоги Т-канальцев. Кроме того, в саркоплазме предсердных кардиомиоцитов у полюсов ядер локализуются специфические предсердные гранулы, состоящие из гликопротеиновых комплексов. Выделяясь из кардиомиоцитов в кровь предсердий, эти вещества влияют на уровень давления крови в сердце и сосудах, а также препятствуют образованию тромбов в предсердиях. Следовательно, предсердные кардиомиоциты, кроме сократительной, обладают и секреторной функцией. В желудочковых кардиомиоцитах более выражены сократительные элементы, а секреторные гранулы отсутствуют.
Вторая разновидность кардиомиоцитов - атипичные кардиомиоциты образуют проводящую систему сердца, состоящую из:
Синусо-предсердный узел;
Предсердно-желудочковый узел;
Предсердно-желудочковый пучок (пучок Гиса), ствол, правую и левую ножки;
Концевые разветвления ножек - волокна Пуркинье.
Атипичные кардиомиоциты обеспечивают генерирование биопотенциалов, их проведение и передачу на сократительные кардиомиоциты.
По своей морфологии атипичные кардиомиоциты отличаются от типичных рядом особенностей:
Они крупнее (длина 100 мкм, толщина 50 мкм);
В цитоплазме содержимся мало миофибрилл, которые расположены неупорядочено и потому атипичные кардиомиоциты не имеют поперечной исчерченности;
Плазмолемма не образует Т-канальцев;
Во вставочных дисках между этими клетками отсутствуют десмосомы и щелевидные контакты.
Атипичные кардиомиоциты различных отделов проводящей системы отличаются между собой по структуре и функциям и подразделяются на три основные разновидности:
· Р-клетки (пейсмекеры) - водители ритма (I типа);
· переходные клетки (II типа);
· клетки пучка Гиса и волокон Пуркинье (III тип).
Клетки I типа (Р-клетки) составляют основу синусо-предсердного узла, а также в небольшом количестве содержатся в атриовентрикулярном узле. Эти клетки способны самостоятельно генерировать с определенной частотой биопотенциалы и передавать их на переходные клетки (II типа), а последние передают импульсы на клетки III типа, от которых биопотенциалы передаются на сократительные кардиомиоциты.
Источники развития кардиомиоцитов - миоэпителиальные пластинки, представляющие собой определенные участки висцеральных листков спланхнотома, а конкретнее из целомического эпителия этих участков.
Гладкая мышечная ткань
Состоит из одноядерных клеток - миоцитов веретеновидной формы длиной 20 - 500 мкм. Их цитоплазма в световом микроскопе выглядит однородно, без поперечной исчерченности. Входит в состав стенок внутренних органов: кровеносных и лимфатических сосудов, мочевыводящих путей, пищеварительного тракта.(сокращение стенок желудка и кишечника)
Фибриллы сократительных белков (миофибриллы), расположенные в их цитоплазме, не имеют той жесткой структурной организации, которая характерна для рассмотренных выше двух других типов волокон. Гладкомышечные волокна имеют удлиненную веретеновидную форму с заостренными концами и центрально расположенным ядром. Гладкомышечные клетки могут образовывать во внутренних органах пласты или тяжи большой протяженности, объединенные соединительнотканными прослойками и пронизанные сосудами и нервами. Работа гладких мышц, как и сердечной, находится под контролем вегетативной нервной системы, и потому они являются непроизвольными. В функциональном отношении они отличаются от других типов мышц тем, что способны осуществлять относительно медленные движения и длительно поддерживать тоническое сокращение. Ритмические сокращения гладких мышц стенок желудка, кишок, мочевого или желчного пузыря обеспечивают перемещение содержимого этих полых органов. Яркий пример – перистальтические движения кишечника, способствующие проталкиванию пищевого комка. Функционирование сфинктеров полых органов непосредственно связано со способностью гладкой мускулатуры к длительным тоническим сокращениям; именно это позволяет надолго перекрывать выход содержимого таких органов, обеспечивая, например, накопление желчи в желчном пузыре. Тонус мышечного слоя стенок артерий определяет величину их просвета и тем самым уровень кровяного давления. При гипертонической болезни (гипертензии) повышенный тонус гладких мышц в стенках малых артерий и артериол приводит к значительному сужению их просвета, повышая сопротивление току крови. Аналогичная картина наблюдается при бронхиальной астме: в ответ на некоторые внешние или внутренние факторы резко возрастает тонус гладких мышц в стенках малых бронхов, вследствие чего просвет бронхов быстро сужается, нарушается выдох и возникает дыхательный спазм.